Carbon Balance and Streamflow at a Small Catchment Scale 10 Years after the Severe Natural Disturbance in the Tatra Mts, Slovakia

Abstract
:1. Introduction
2. Material and Methods
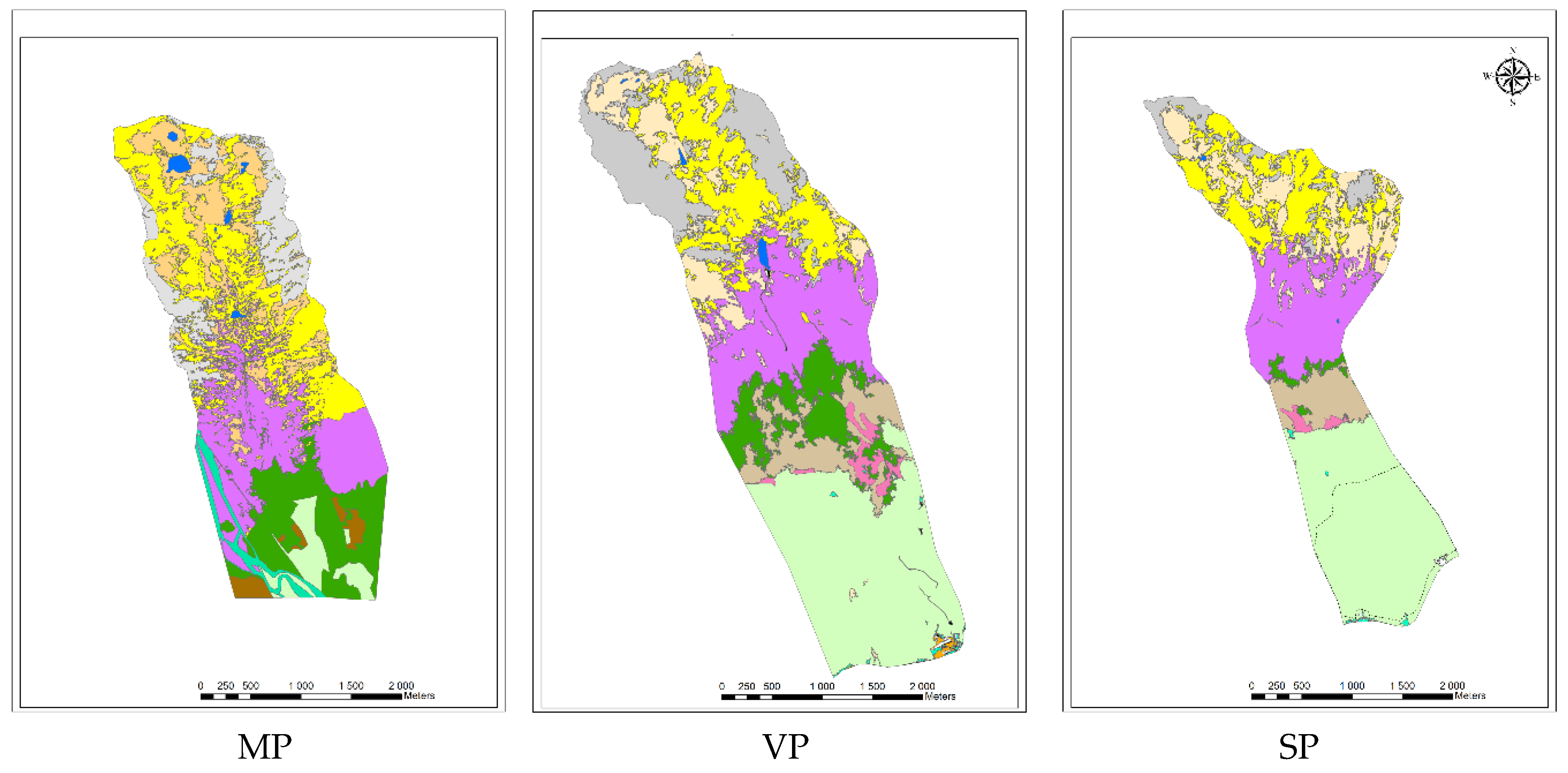
2.1. Study Sites
2.2. Meteorological and Hydrological Data
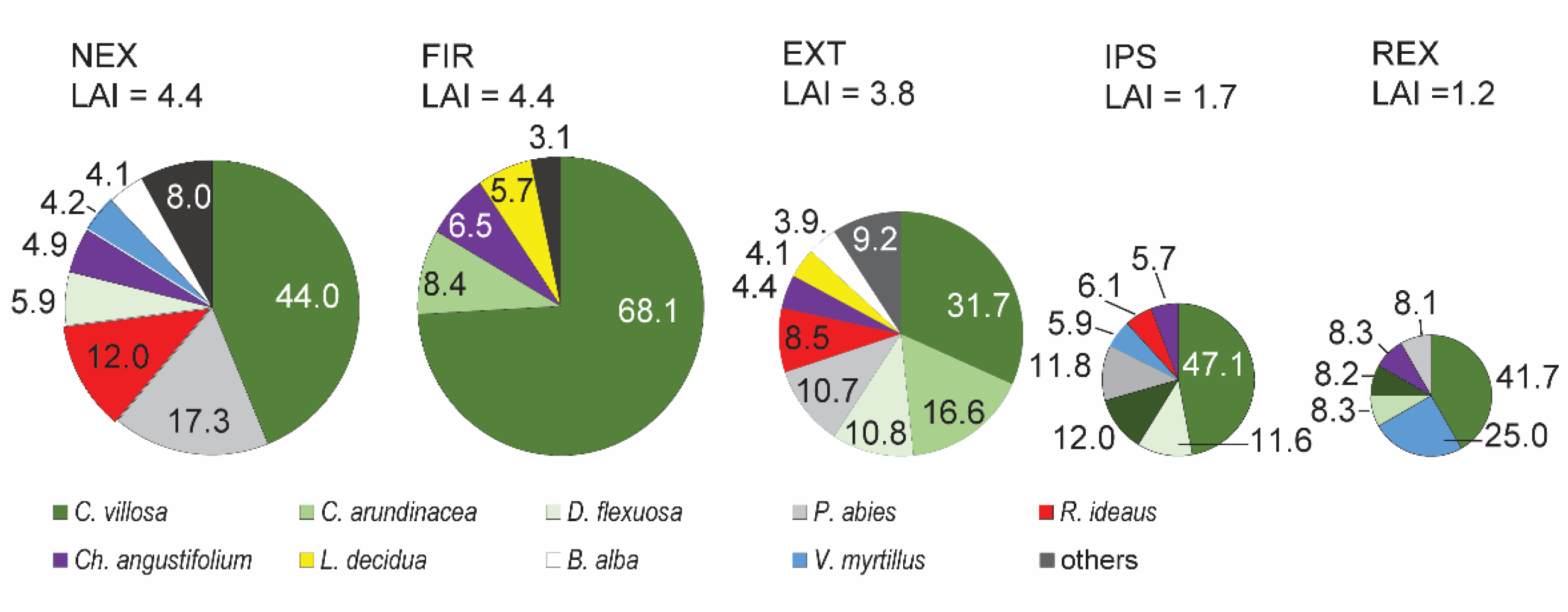
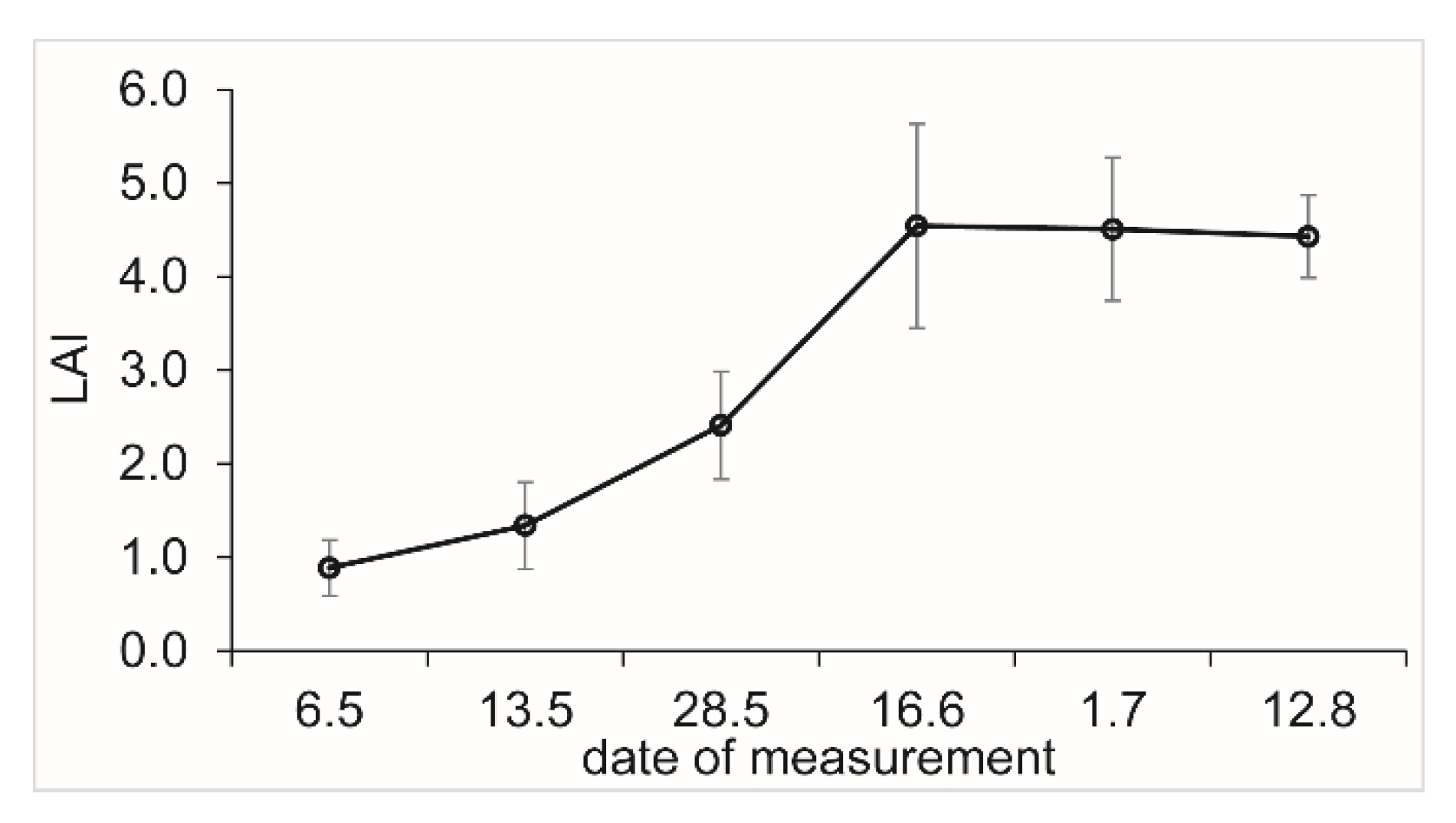
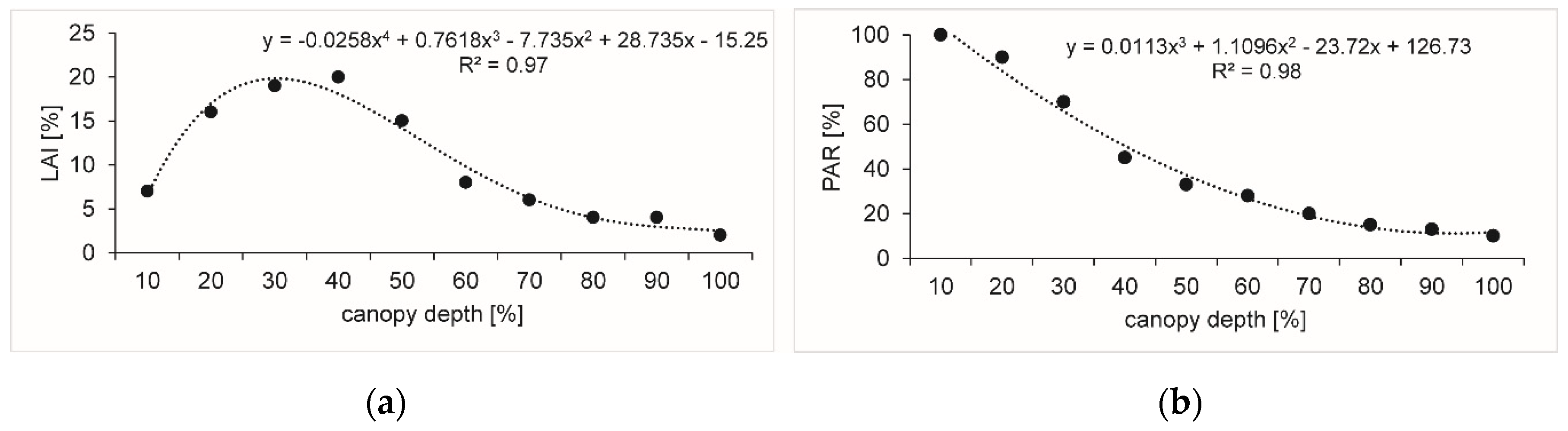
2.3. Vegetation and Phenology
2.4. Carbon Balance
2.4.1. Assimilation—GPP
2.4.2. Ecosystem Respiration—RE
2.4.3. Flux Data Validation
2.5. Streamflow Characteristics
3. Results
3.1. Weather, Vegetation and Phenology
3.2. Carbon Fluxes
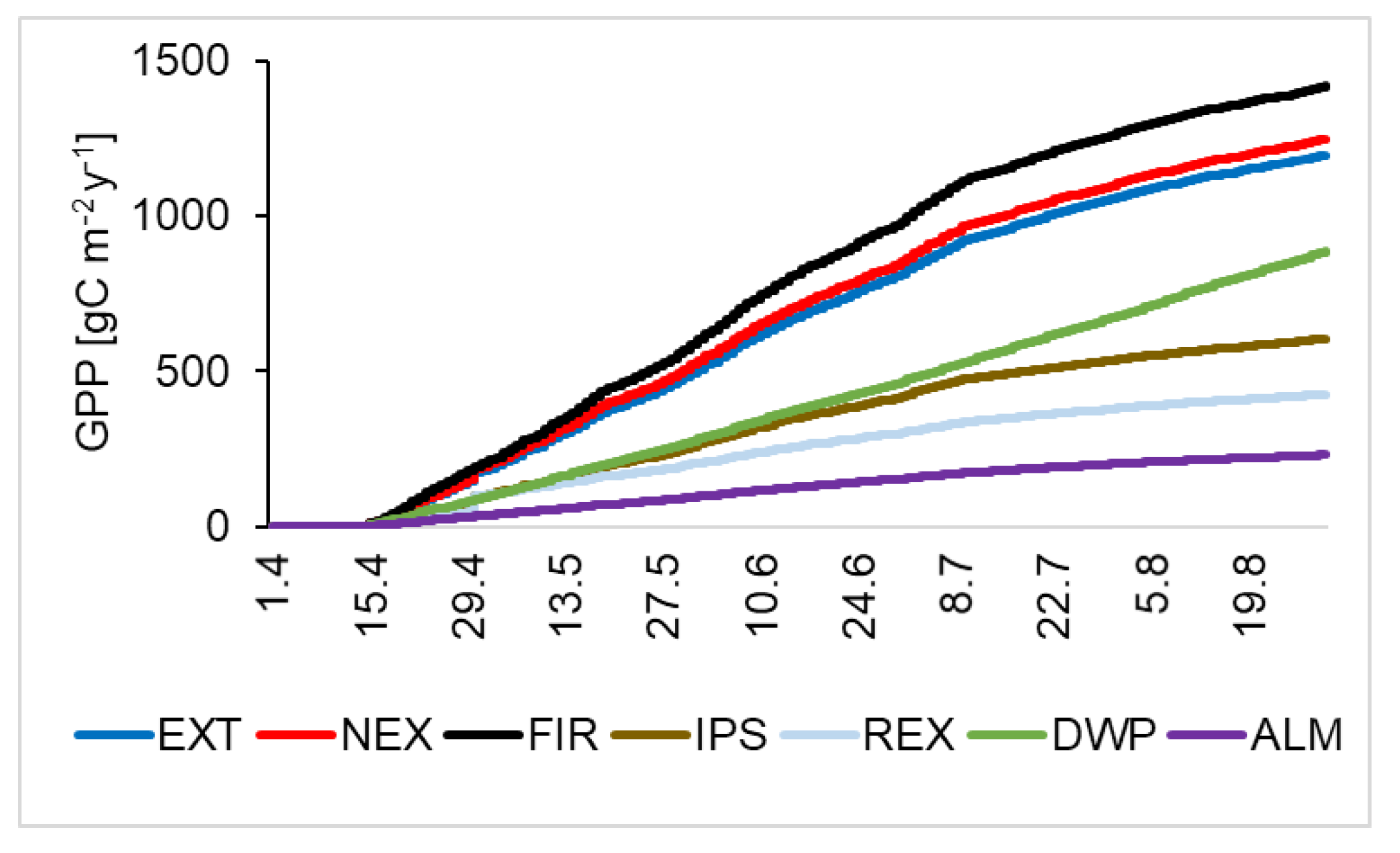
3.2.1. Assimilation (GPP)
3.2.2. Ecosystem (RE) and Component Respiration
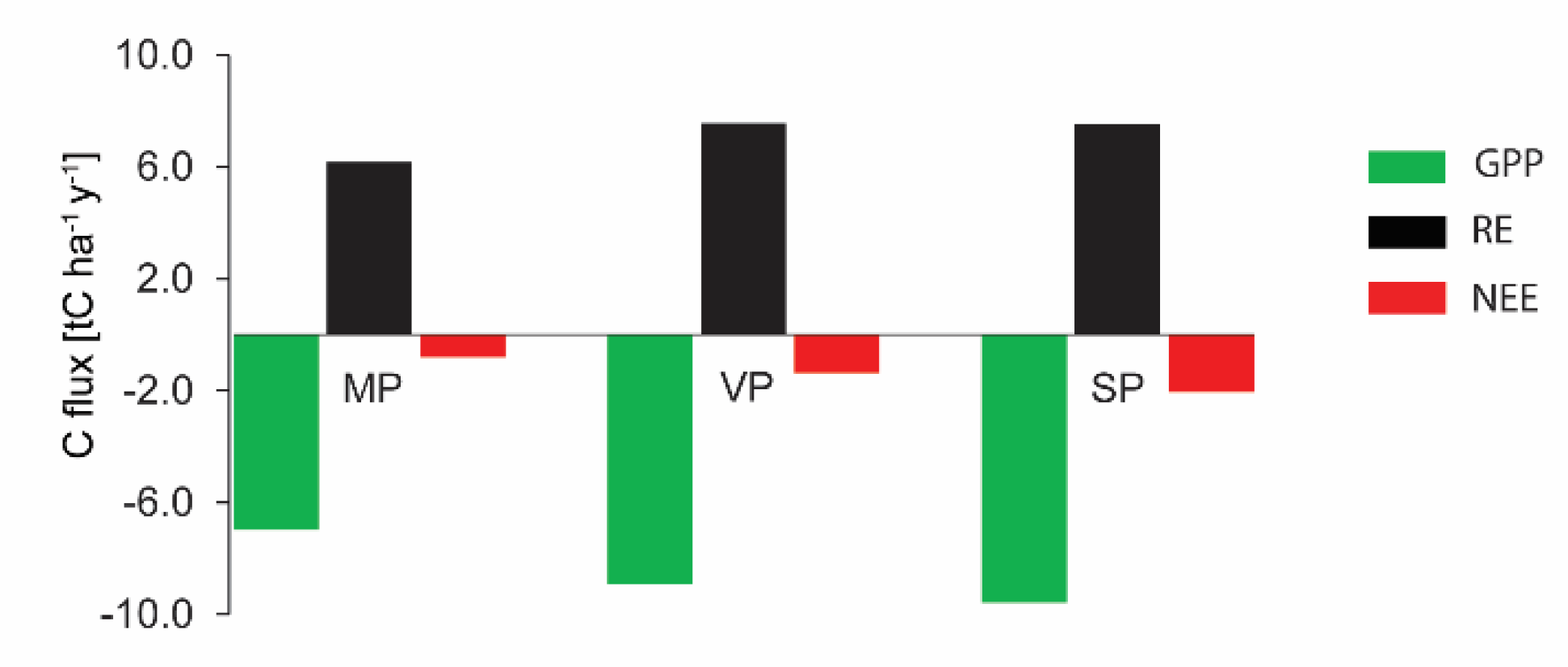
3.2.3. Carbon Balance (NEE)
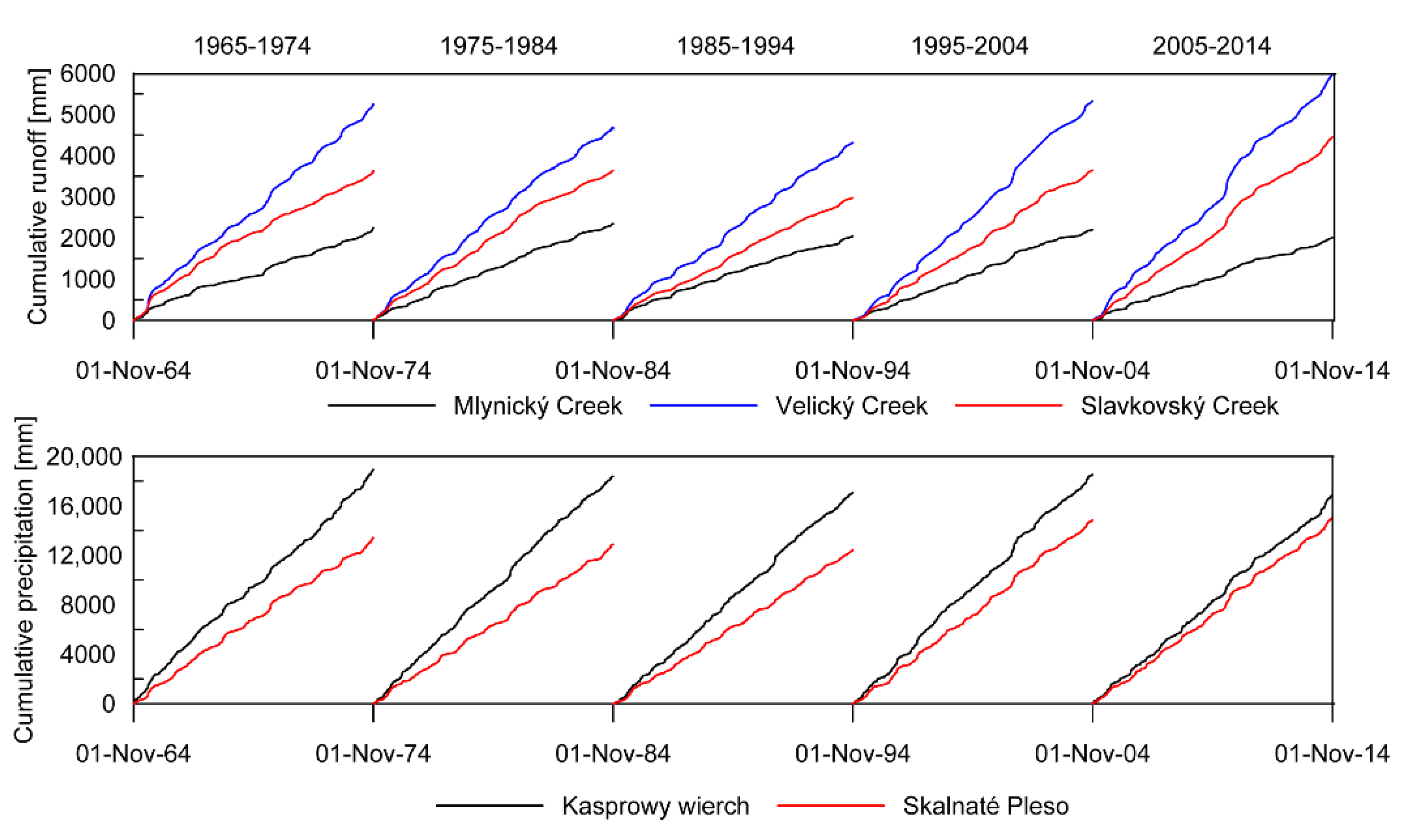
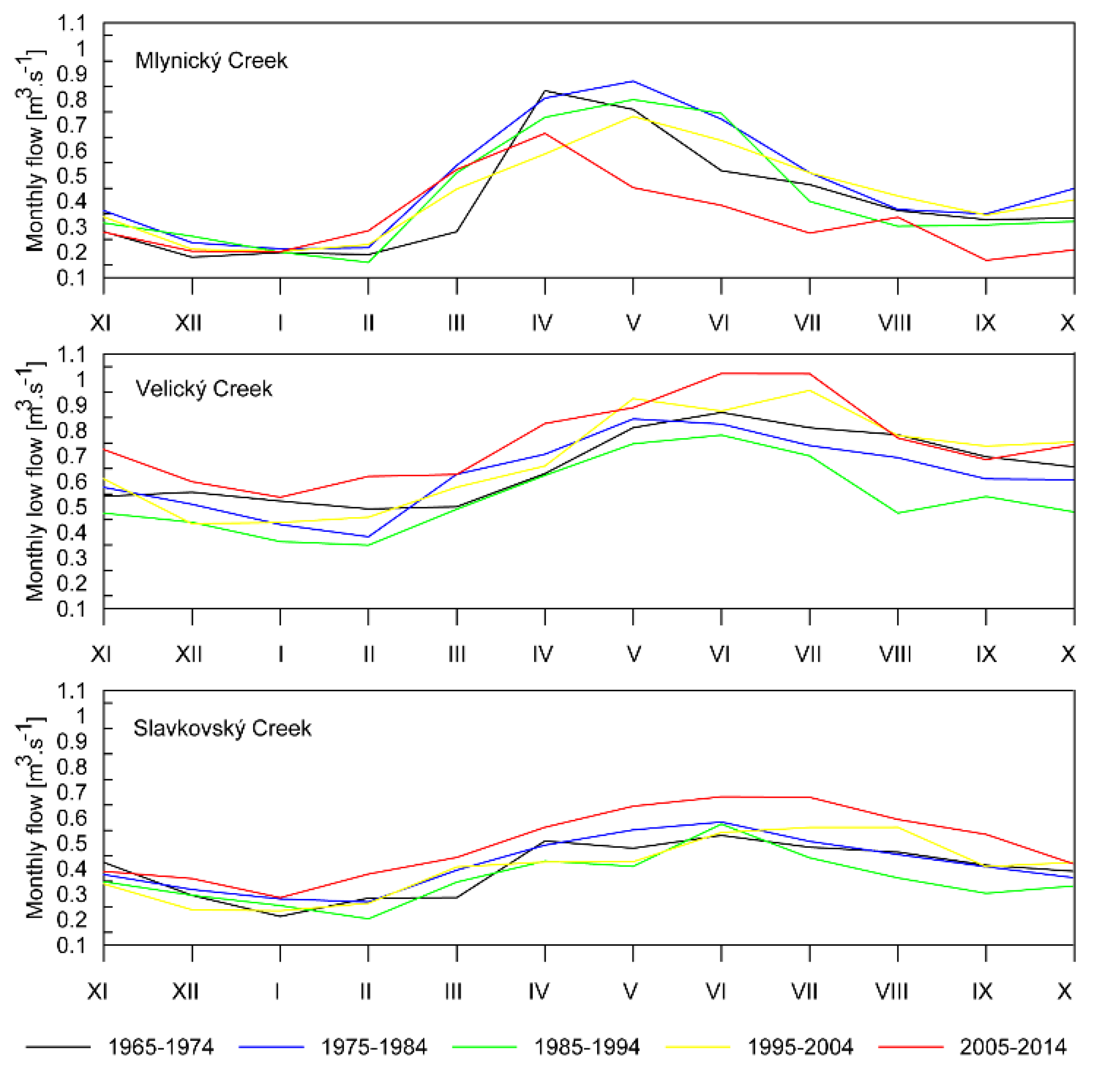
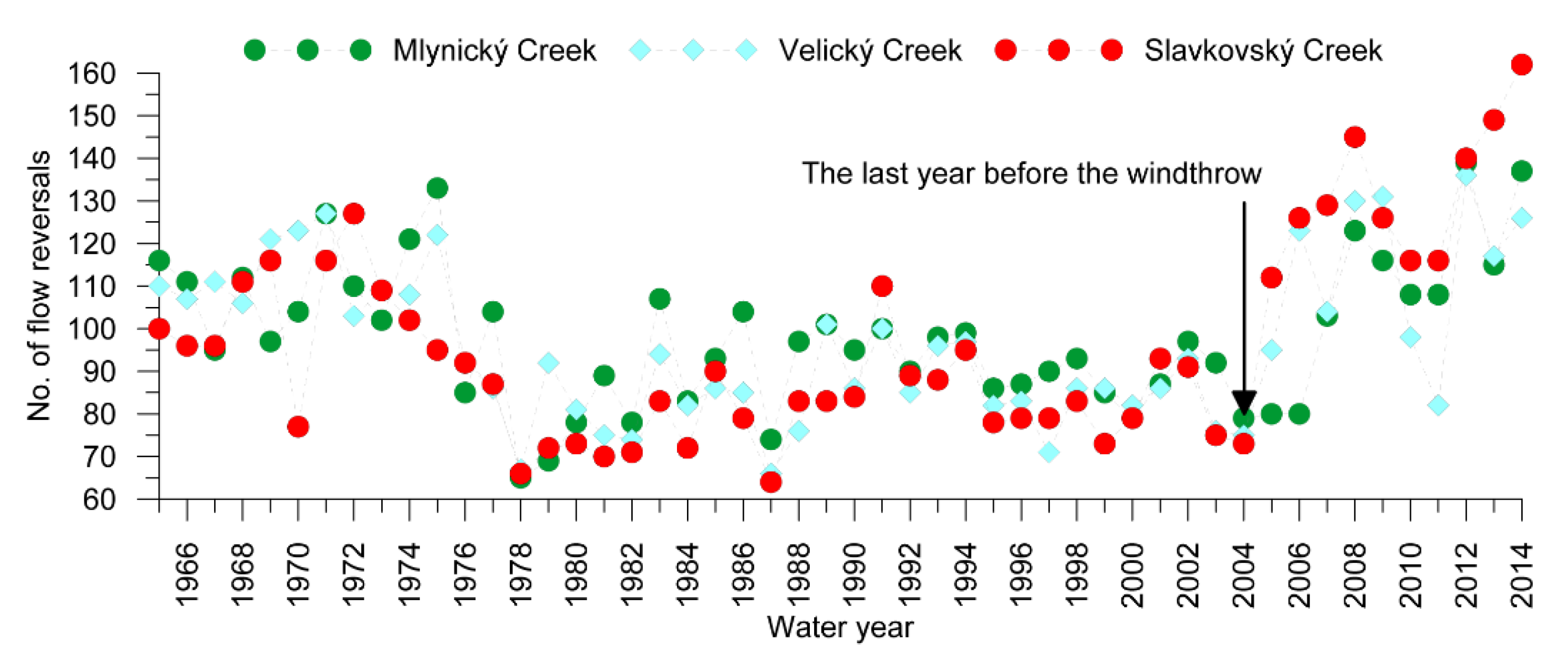
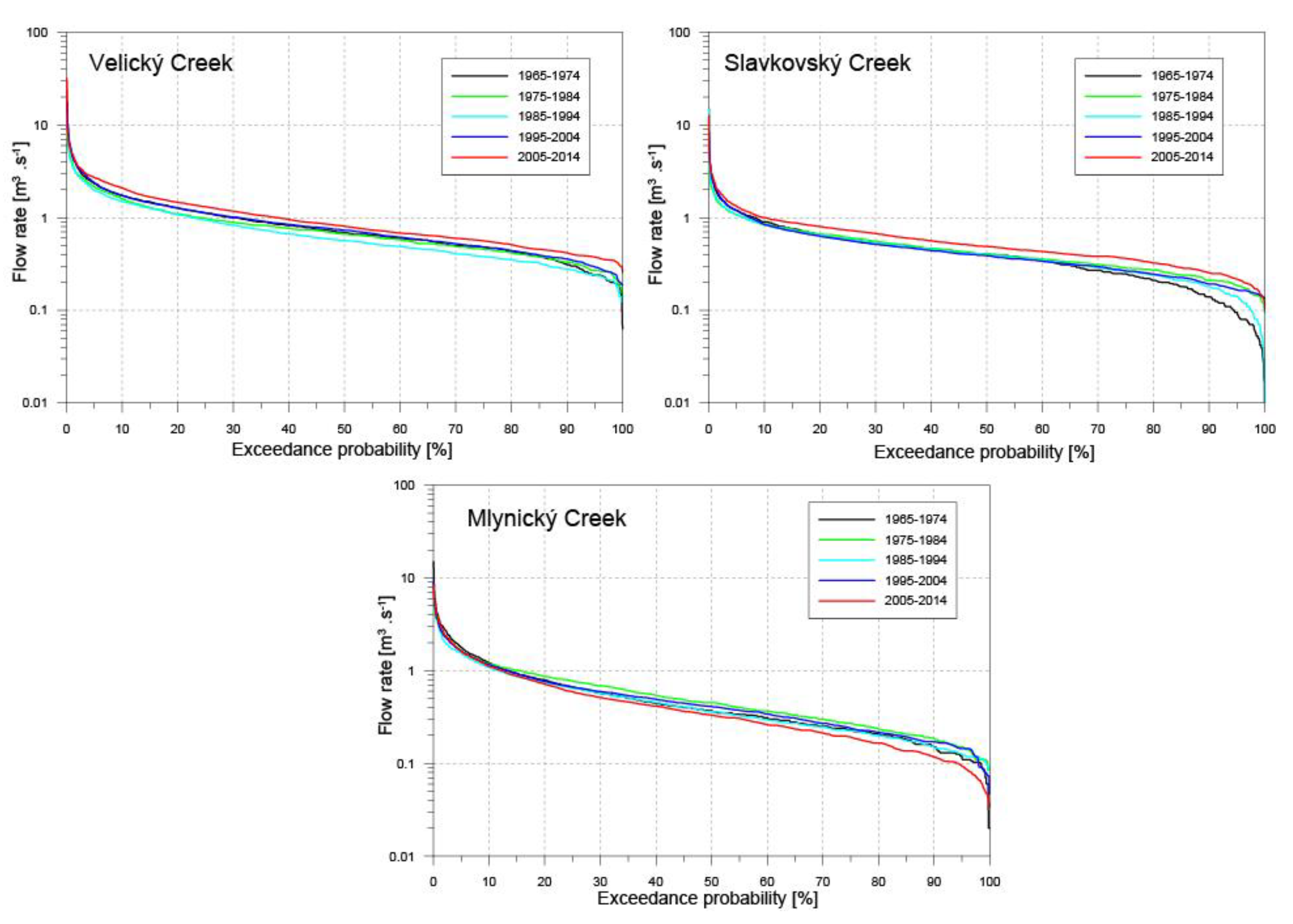
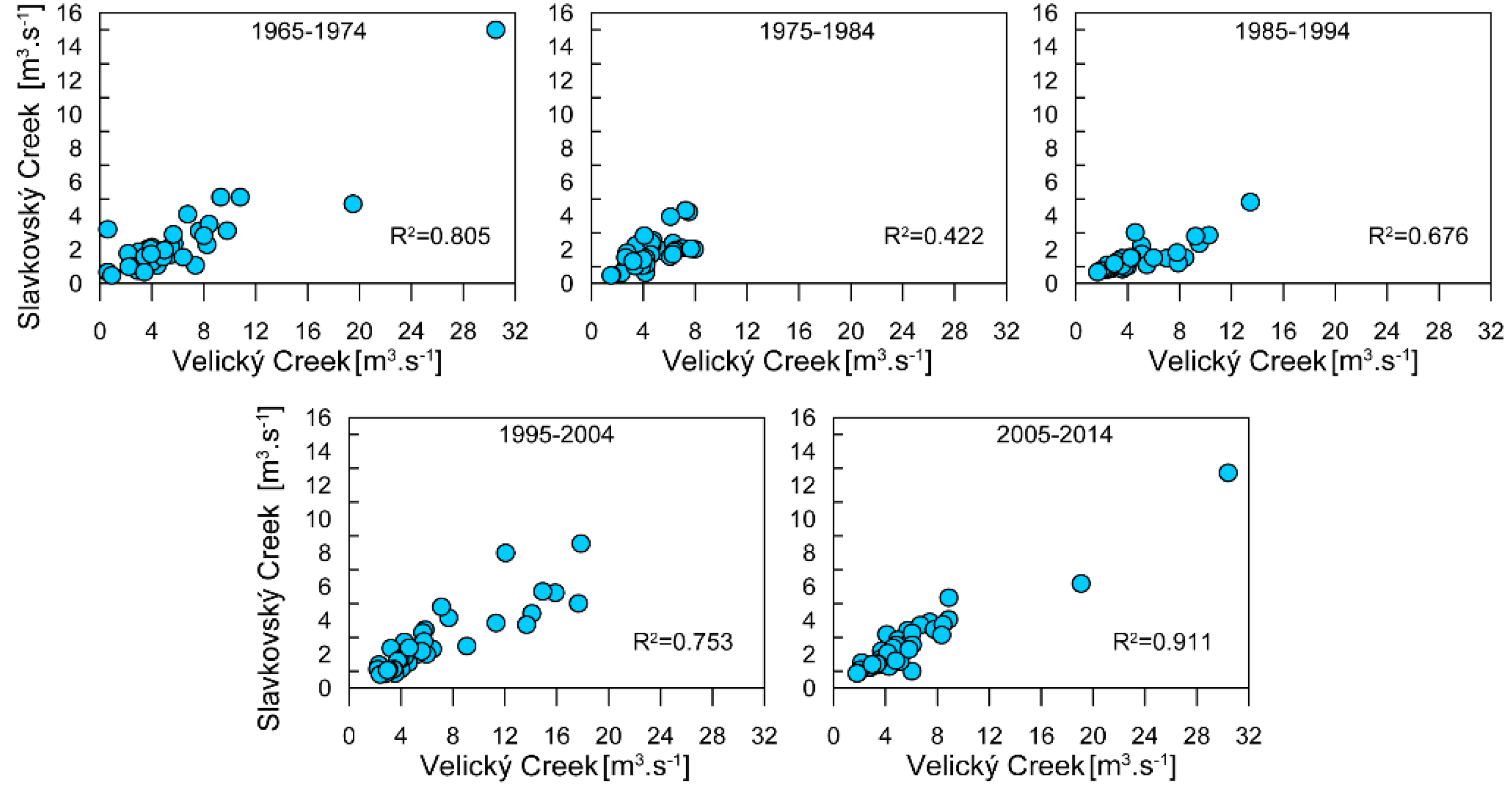
3.3. Streamflow Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thom, D.; Seidl, R. Natural disturbance impacts on ecosystem services and biodiversity in temperate and boreal forests. Biol. Rev. 2015, 91, 760–781. [Google Scholar] [CrossRef] [PubMed]
- Senf, C.; Seidl, R. Mapping the coupled human and natural disturbance regimes of Europe’s forests. BioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Seidl, R.; Schelhaas, M.J.; Lexer, M.J. Unraveling the drivers of intensifying forest disturbance regimes in Europe. Glob. Chang. Biol. 2011, 17, 2842–2852. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Kane, J.M.; Anderegg, L.D.L. Consequences of widespread tree mortality triggered by drought and temperature stress. Nat. Clim. Chang. 2013, 3, 30–36. [Google Scholar] [CrossRef]
- Malhi, Y.; Franklin, J.; Seddon, N.; Solan, M.; Turner, M.G.; Field, C.B.; Knowlton, N. Climate change and ecosystems: Threats, opportunities and solutions. Philos. Trans. R. Soc. B 2020, 375. [Google Scholar] [CrossRef] [Green Version]
- Weiskopf, S.R.; Rubenstein, M.A.; Crozier, L.G.; Gaichas, S.; Griffis, R.; Halofsky, J.E.; Hyde, K.J.W.; Morelli, T.L.; Morisette, J.T.; Muñoz, R.C.; et al. Climate change effects on biodiversity, ecosystems, ecosystem services, and natural resource management in the United States. Sci. Total Environ. 2020, 733. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2013. [Google Scholar]
- Perdrial, J.; Brooks, P.D.; Swetnam, T.; Lohse, K.A.; Rasmussen, C.; Litvak, M.; Harpold, A.A.; Zapata-Rios, X.; Broxton, P.; Mitra, B.; et al. A net ecosystem carbon budget for snow dominated forested headwater catchments: Linking water and carbon fluxes to critical zone carbon storage. Biogeochemistry 2018, 138, 225–243. [Google Scholar] [CrossRef]
- Bellassen, V.; Luyssaert, S. Managing forests in uncertain times. Nature 2014, 506, 153–155. [Google Scholar] [CrossRef] [Green Version]
- Nabuurs, G.; Schelhaas, M. Carbon profiles of typical forest types across Europe assessed with CO2FIX. Ecol. Indic 2002, 1, 213–223. [Google Scholar] [CrossRef]
- Harmon, M.E.; Bond-Lamberty, B.; Tang, J.; Vargas, R. Heterotrophic respiration in disturbed forests: A review with examples from North America. J. Geophys. Res. Biogeosci. 2011, 116, 1–17. [Google Scholar] [CrossRef]
- Amiro, B.D.; Barr, A.G.; Barr, J.G.; Black, T.A.; Bracho, R.; Brown, M.; Chen, J.; Clark, K.L.; Davis, K.J.; Desai, A.R.; et al. Ecosystem carbon dioxide fluxes after disturbance in forests of North America. J. Geophys. Res. 2010, 115, G00K02. [Google Scholar] [CrossRef]
- Kurz, W.A.; Dymond, C.G.; Stinson, G.J.; Rampley, G.J.; Neilson, E.T.; Carroll, A.L.; Ebata, T.; Safranyik, L. Mountain pine beetle and forest carbon feedback to climate change. Nature 2008, 452, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Niu, S.; Fu, Z.; Luo, Y.; Stoy, P.C.; Keenan, T.F.; Poulter, B.; Zhang, L.; Piao, S.; Zhou, X.; Zheng, H.; et al. Interannual variability of ecosystem carbon exchange: From observation to prediction. Glob. Ecol. Biogeogr. 2017, 26, 1225–1237. [Google Scholar] [CrossRef]
- Calderón-Loor, M.; Cuesta, F.; Pinto, E.; Gosling, W.D. Carbon sequestration rates indicate ecosystem recovery following human disturbance in the equatorial Andes. PLoS ONE 2020, 15, e0230612. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, W.R.L.; Martinez-Vilalta, J.; Cailleret, M.; Camarero, J.J.; Ewers, B.E.; Galbraith, D.; Gessler, A.; Grote, R.; Huang, C.; Levick, S.R.; et al. When a Tree Dies in the Forest: Scaling Climate-Driven Tree Mortality to Ecosystem Water and Carbon Fluxes. Ecosystems 2016, 19, 1133–1147. [Google Scholar] [CrossRef] [Green Version]
- Levy-Varon, J.H.; Schuster, W.S.F.; Griffin, K.L. Rapid rebound of soil respiration following partial stand disturbance by tree girdling in a temperate deciduous forest. Oecologia 2014, 174, 1415–1424. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.; Rammer, W.; Verkerk, P. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Chang. 2014, 4, 806–810. [Google Scholar] [CrossRef] [Green Version]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Senevirante, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef]
- Hlásny, T.; Barcza, Z.; Fabrika, M.; Balász, B.; Churkina, G.; Pajtík, J.; Sedmák, R.; Turčáni, M. Climate change impacts on growth and carbon balance of forests in Central Europe. Clim. Res. 2011, 47, 219–236. [Google Scholar] [CrossRef] [Green Version]
- Pugh, T.A.M.; Arneth, A.; Kautz, M.; Poulter, B.; Smith, B. Important role of forest disturbances in the global biomass turnover and carbon sinks. Nat. Geosci. 2019, 12, 730–735. [Google Scholar] [CrossRef]
- Klein, D.; Höllerl, S.; Blaschke, M.; Schulz, C. The contribution of managed and unmanaged forests to climate change mitigation-A model approach at stand level for the main tree species in Bavaria. Forests 2013, 4, 43–69. [Google Scholar] [CrossRef]
- Moore, P.T.; DeRose, R.J.; Long, J.N.; van Miegroet, H. Using Silviculture to Influence Carbon Sequestration in Southern Appalachian Spruce-Fir Forests. Forests 2012, 3, 300–316. [Google Scholar] [CrossRef]
- Baldocchi, D.D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: Past, present and future. Glob. Chang. Biol. 2003, 9, 479–492. [Google Scholar] [CrossRef] [Green Version]
- Noe, S.M.; Kimmel, V.; Hüve, K.; Copolovici, L.; Portillo-Estrada, M.; Püttsepp, Ü.; Jõgiste, K.; Niinemets, Ü.; Hörtnagl, L.; Wohlfahrt, G. Ecosystem-scale biosphere-atmosphere interactions of a hemiboreal mixed forest stand at Järvselja, Estonia. For. Ecol. Manag. 2011, 262, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Lindauer, M.; Schmid, H.P.; Grote, R.; Mauder, M.; Steinbrecher, R.; Wolpert, B. Net ecosystem exchange over a non-cleared wind-throw-disturbed upland spruce forest—Measurements and simulations. Agric. For. Meteorol. 2014, 197, 219–234. [Google Scholar] [CrossRef]
- Cleary, M.B.; Naithani, K.J.; Ewers, B.E.; Pendall, E. Upscaling CO2 fluxes using leaf, soil and chamber measurements across successional growth stages in a sagebrush steppe ecosystem. J. Arid. Environ. 2015, 121, 43–51. [Google Scholar] [CrossRef]
- Wang, K.; Liu, C.; Zheng, X.; Pihlatie, M.; Li, B.; Haapanala, S.; Vesala, T.; Liu, H.; Wang, Y.; Liu, G.; et al. Comparison between eddy covariance and automatic chamber techniques for measuring net ecosystem exchange of carbon dioxide in cotton and wheat fields. Biogeosciences 2013, 10, 6865–6877. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Chen, J.M.; Liu, J.; Black, T.A.; Croft, H.; Staebler, R.; McCaughey, H. Comparison of big-leaf, two-big-leaf, and two-leaf upscaling schemes for evapotranspiration estimation using coupled carbon-water modeling. J. Geophys. Res. Biogeosci. 2018, 123, 207–225. [Google Scholar] [CrossRef]
- Kostka, Z.; Holko, L. Role of forest in hydrological cycle—Forest and runoff. Meteorol. J. 2006, 9, 143–148. [Google Scholar]
- Andreassian, V. Waters and forests: From historical controversy to scientific debate. J. Hydrol. 2004, 291, 1–27. [Google Scholar] [CrossRef]
- Ben-Hur, M.; Fernandez, C.; Sakkola, S.; Santamarta-Cerezal, J.C. Overland flow, soil erosion and stream water quality in forest under diferent pertubations and climate conditions. In Forest Management and Water Cycle; Bredemeier, M., Cohen, S., Godbold, D., Lode, E., Pichler, V., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 263–289. [Google Scholar]
- Bartík, M.; Holko, L.; Jančo, M.; Škvarenina, J.; Danko, M.; Kostka, Z. Influence of mountain spruce forest dieback on snow accumulation and melt. J. Hydrol. Hydromech. 2019, 67, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Bosch, J.M.; Hewlett, J.D. A review of catchment ex-periments to determine the effect of vegetation changes on water yield and evapotranspiration. J. Hydrol. 1982, 55, 3–23. [Google Scholar] [CrossRef]
- Best, A.; Zhang, L.; McMahon, T.; Western, A.; Vertessy, R. A Critical Review of Paired Catchment Studies with Reference to Seasonal Flows and Climatic Variability. In CSIRO Land and Water Technical Report 25/03; Murray Darling Basin Commission: Canberra, Australia, 2003; p. 56. ISBN 1 876 830 57 3. [Google Scholar]
- Jones, J.A.; Post, D.A. Seasonal and successional streamflow response to forest cutting and regrowth in the northwestern and eastern United States. Water Res. 2004, 40, W05203. [Google Scholar] [CrossRef]
- Stednick, J.D. Monitoring the effects of timber harvest on annual water yield. J. Hydrol. 1996, 176, 79–95. [Google Scholar] [CrossRef]
- Alila, Y.; Kuraś, P.K.; Schnorbus, M.; Hudson, R. Forests and floods: A new paradigm sheds light on age-old controversies. Water Resour. Res. 2009, 45, W08416. [Google Scholar] [CrossRef]
- The Nature Conservancy. Indicators of Hydrologic Alteration Version 7.1 User’s Manual. Nat. Conserv. 2009. Available online: https://www.conservationgateway.org/Files/Pages/indicators-hydrologic-altaspx47.aspx (accessed on 1 March 2014).
- Olden, J.D.; Poff, N.L. Redundancy and the choice of hydrologic indices for characterizing streamflow regimes. River Res. Appl. 2003, 19, 101–121. [Google Scholar] [CrossRef]
- Gao, Y.; Vogel, R.M.; Kroll, C.N.; Olden, J.D. Development of representative indicators of hydrologic alteration. J. Hydrol. 2009, 374, 136–147. [Google Scholar] [CrossRef]
- Vogel, R.M.; Sieber, J.; Archfield, S.A.; Smith, M.P.; Apse, C.D.; Huber-Lee, A. Relations among storage, yield, and instream flow. Water Resour. Res. 2007, 43, W05403. [Google Scholar] [CrossRef]
- Holeksa, J.; Zielonka, T.; Zywiec, M.; Fleischer, P. Identifying the disturbance history over a large area of larch-spruce mountain forest in Central Europe. For. Ecol. Manag. 2016, 361, 318–327. [Google Scholar] [CrossRef]
- Fleischer, P.; Homolová, Z. Long-term ecological research in larch-spruce forest community after natural disturbances in the Tatra Mts. Lesn. Cas. For. J. 2011, 57, 237–250. [Google Scholar]
- Holko, L.; Fleischer, P.; Novák, V.; Kostka, Z.; Bičárová, S.; Novák, J. Hydrological Effects of a Large Scale Windfall Degradation in the High Tatra Mountains, Slovakia. In Management of Mountain Watersheds; Křeček, J., Haigh, M., Hofer, T., Kubin, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 164–179. [Google Scholar]
- Holko, L.; Hlavatá, H.; Kostka, Z.; Novák, J. Hydrological regimes of small catchments in the High Tatra Mountains before and after extraordinary wind-induced deforestation. Folia Geogr. Ser. Geogr. Phys. 2009, 15, 33–44. [Google Scholar]
- Holko, L.; Kostka, Z.; Novák, J. Estimation of groundwater recharge, water balance of small catchments in the High Tatra Mountains in hydrological year 2008. In Sustainable Development and Bioclimate: Reviewed Conference Proceedings; Pribullová, A., Bičárová, S., Eds.; Geophysical Institute of the Slovak Academy of Sciences: Bratislava, Slovakia, 2009; pp. 93–94. [Google Scholar]
- Holko, L.; Škoda, P. Assessment of runoff changes in selected catchments of the High Tatra Mountains ten years after the windthrow. Acta Hydrolog. Slovaca 2016, 17, 43–50. (In Slovak) [Google Scholar]
- Rasband, W.S. Image J. U.S. National Institutes of Health, Bethesda, Maryland, USA. Available online: https://imagej.nih.gov/ij/ (accessed on 30 September 2015).
- Pajtík, J.; Konôpka, B.; Lukac, M. Biomass functions and expansion factors in young Norway spruce (Picea abies [L.] Karst) trees. For. Ecol. Manag. 2008, 256, 1096–1103. [Google Scholar] [CrossRef]
- Pajtík, J.; Konôpka, B.; Bošela, M.; Šebeň, V.; Kaštier, P. Modelling forage potential for red deer: A case study in post-disturbance young stands of rowan. Ann. For. Res. 2015, 58, 91–107. [Google Scholar] [CrossRef] [Green Version]
- Pajtík, J.; Konôpka, B.; Šebeň, V.; Michelčík, P.; Fleischer, P. Biomass allocation of common larch in the first age class in the High Tatra Mts. Stud. Tatra Natl. Park. 2015, 44, 237–249. [Google Scholar]
- Pajtík, J.; Konôpka, B. Quantifying edible biomass on young Salix caprea and Sorbus aucuparia trees for Cervus elaphus: Estimates by regression models. Austrian J. For. Sci. 2015, 132, 61–80. [Google Scholar]
- Fleischer, P.; Fleischer, P., Jr.; Gömöryová, E.; Celer, S. Upscaling carbon fluxes from chamber measurement to the landscape-scale in spruce forest disturbed by windthrow in the Tatra Mts. In Landscape and Landscape Ecology, Proceedings of the 17th int symposium on Landscape Ecology; Halada, L., Bača, A., Boltižiar, M., Eds.; Institute of Landscape Ecology, Slovak Academy of Sciences: Nitra, Slovakia, 2016; pp. 227–235. [Google Scholar]
- Drewitt, G.B.; Black, T.A.; Nesic, Z.; Humphreys, E.R.; Jork, E.M.; Swanson, R.; Ethier, G.J.; Griffis, T.; Morgenstern, K. Measuring forest floor CO2 fluxes in a Douglas-fir forest. Agric. For. Meteorol. 2002, 110, 299–317. [Google Scholar] [CrossRef]
- Fleischer, P.; Fleischer, P., Jr.; Celer, S. Carbon balance in spruce forest affected by wind and insect disturbances in the Tatra Mts: Methodological approach and preliminary results. In Aktuálne Problémy v Ochrane Lesa; Kunca, A., Ed.; National Forest Center: Zvolen, Slovakia, 2013; pp. 113–120. (In Slovak) [Google Scholar]
- Marek, M.V. Carbon in Ecosystems under Changing Climate in Czech Republic; Academia Praha: Prague, Czech Repulic, 2011. (In Czech) [Google Scholar]
- Ryan, M.G.; Lavigne, M.B.; Gower, S.T. Annual carbon cost of autotrophic respiration in boreal forest ecosystems in relation to species and climate. J. Geophys. Res. 1997, 102, 28871–28883. [Google Scholar] [CrossRef] [Green Version]
- Buchmann, N. Biotic and abiotic factors controlling soil respiration rates in Picea abies stands. Soil. Biol. Biochem. 2000, 32, 1625–1635. [Google Scholar] [CrossRef]
- Knohl, A.; Søe, A.R.B.; Kutsch, W.L.; Göckede, M.; Buchmann, N. Representative estimates of soil and ecosystem respiration in an old beech forest. Plant Soil 2008, 302, 189–202. [Google Scholar] [CrossRef] [Green Version]
- Searsy, J.K. Flow-Duration Curves; Geological Survey Water-Supply-Papet 1542-a: Washington, DC, USA, 1959; p. 33. [Google Scholar]
- Curtis, P.S.; Gough, C.M. Forest aging, disturbance and the carbon cycle. N. Phytol. 2018, 219, 1188–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Jose, S.; Hu, Z.; Pang, J.; Hou, L.; Zhang, S. Biometric and eddy covariance methods for examining the carbon balance of a larix principis-rupprechtii forest in the Qinling Mountains, China. Forests 2018, 9, 67. [Google Scholar] [CrossRef] [Green Version]
- Rebane, S.; Jõgiste, K.; Kiviste, A.; Stanturf, J.A.; Metslaid, M. Patterns of carbon sequestration in a young forest ecosystem after clear-cutting. Forests 2020, 11, 126. [Google Scholar] [CrossRef] [Green Version]
- Amiro, B.D.; Barr, A.G.; Black, T.A.; Iwashita, H.; Kljun, N.; McCaughey, H.; Morgenstern, K.; Murayama, S.; Nesic, Z.; Orchansky, A.L.; et al. Carbon, energy and water fluxes at mature and disturbed forest sites, Saskatchewan, Canada. Agric. For. Meteorol. 2006, 136, 237–251. [Google Scholar] [CrossRef]
- Lindroth, A.; Lagergren, F.; Grelle, A.; Klemedtsson, L.; Langvall, O.; Weslien, P.; Tuulik, J. Storms can cause Europe-wide reduction in forest carbon sink. Glob. Chang. Biol. 2009, 15, 346–355. [Google Scholar] [CrossRef]
- Giorgi, S.; Fleischer, P.; Gioli, B.; Kolle, O.; Manca, G.; Matese, A.; Zaldei, A.; Ziegler, W.; Cescatti, A.; Miglietta, F.; et al. Carbon Dioxide Fluxes of a Recent Large Windthrow in the High Tatra. Abstract of CEIP Conference. Sissi-Lassithi, Crete. 2006. Available online: http://carboeurope.org/ceip/conference/abstacts/VU_poster/Giorgi) (accessed on 1 November 2006).
- Lindroth, A.; Grelle, A.; Morén, A.S. Long-term measurements of boreal forest carbon balance reveal large temperature sensitivity. Glob. Chang. Biol. 1998, 4, 443–450. [Google Scholar] [CrossRef]
- Zhu, K.; Zhang, J.; Niu, S.; Chu, C.; Luo, Y. Limits to growrh of forest biomass carbon sink under climate change. Nat. Commun. 2020. [Google Scholar] [CrossRef] [Green Version]
- Michalová, Z.; Morrissey, R.C.; Wohlgemuth, T.; Bače, R.; Fleischer, P.; Svoboda, M. Salvage-Logging after Windstorm Leads to Structural and Functional Homogenization of Understory Layer and Delayed Spruce Tree Recovery in Tatra Mts., Slovakia. Forests 2017, 8, 88. [Google Scholar] [CrossRef] [Green Version]
- Pumpanen, J.; Westman, C.J.; Ilvesniemi, H. Soil CO2 efflux from a podzolic forest soil before and after forest clear-cutting and site preparation. Boreal Environ. Res. 2004, 9, 199–212. [Google Scholar]
- Koster, K.; Puttsepp, U.; Pumpanen, J. Comparison of soil CO2 flux between uncleared and cleared windthrow areas in Estonia and Latvia. For. Ecol. Manag. 2011, 262, 65–70. [Google Scholar] [CrossRef]
- Morehouse, K.; Johns, T.; Kaye, J.; Kaye, M. Carbon and nitrogen cycling immediately following bark beetle outbreaks in southwestern ponderosa pine forests. For. Ecol. Manag. 2008, 255, 2698–2708. [Google Scholar] [CrossRef]
- Fleischer, P.; Koreň, M. Selected forest soil properties after the 2004 windfall in the Tatra Mts. In Sustainable Development and Bioclimate; Pribullová, A., Bičarová, S., Eds.; Geophysical Institute of the Slovak Academy of Sciences: Bratislava, Slovakia, 2009; pp. 77–78. [Google Scholar]
- Moore, D.J.P.; Trahan, N.A.; Wilkes, P.; Quaife, T.; Stephens, B.B.; Elder, K.; Desai, A.R.; Negron, J.; Monson, R.K. Persistent reduced ecosystem respiration after insect disturbance in high elevation forests. Ecol. Lett. 2013, 16, 731–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagedorn, F.; Mulder, J.; Jandl, R. Mountain soils under a changing climate and land-use. Biogeochemistry 2010, 97, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Don, A.; Bärwolff, M.; Kalbitz, K.; Andruschkewitsch, R.; Jungkunst, H.F.; Schulze, E.D. No rapid soil carbon loss after a windthrow event in the High Tatra. For. Ecol. Manag. 2012, 276, 239–246. [Google Scholar] [CrossRef]
- Valentini, R. EUROFLUX: An integrated network for studying the long-term responses of biospheric exchanges of carbon, water, and energy of European forests. Fluxes Carbon. Water Energy Eur. For. 2003, 163, 1–8. [Google Scholar]
- Grünwald, T.; Bernhofer, C. A decade of carbon, water and energy flux measurements of an old spruce forest at the Anchor Station Tharandt. Tellus Ser. B Chem. Phys. Meteorol. 2007, 59, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Etzold, S.; Ruehr, N.K.; Zweifel, R.; Dobbertin, M.; Zingg, A.; Pluess, P.; Häsler, R.; Eugster, W.; Buchmann, N. The Carbon Balance of Two Contrasting Mountain Forest Ecosystems in Switzerland: Similar Annual Trends, but Seasonal Differences. Ecosystems 2011, 14, 1289–1309. [Google Scholar] [CrossRef]
- Taylor, A.R.; Seedre, M.; Brassard, B.W.; Chen, H.Y.H. Decline in Net Ecosystem Productivity Following Canopy Transition to Late-Succession Forests. Ecosystems 2014, 17, 778–791. [Google Scholar] [CrossRef]
- Luyssaert, S.; Schulze, E.D.; Börner, A.; Knohl, A.; Hessenmöller, D.; Law, B.E.; Ciais, P.; Grace, J. Old-growth forests as global carbon sinks. Nature 2008, 455, 213–215. [Google Scholar] [CrossRef]
- Yi, C.; Li, R.; Wolbeck, J.; Xu, X.; Nilsson, M.; Aires, L.; Albertson, J.D.; Ammann, C.; Arain, M.A.; De Araujo, A.C.; et al. Climate control of terrestrial carbon exchange across biomes and continents. Environ. Res. Lett. 2010, 5. [Google Scholar] [CrossRef]
- Kulmala, L.; Pumpane, J.; Hari, P.; Vesala, T. Photosynthesis of ground vegetation in different aged pine forests: Effect of environmental factors predicted with a process-based model. J. Veg. Sci. 2011, 22, 96–110. [Google Scholar] [CrossRef]
- Górnik, M.; Holko, L.; Pociask-Karteczka, J.; Bičárová, S. Varibility of precipitation and runoff in the entire High Tatra Mountains in the period 1961–2010. Prace Geograficzne 2017, 151, 53–74. [Google Scholar] [CrossRef] [Green Version]
- Holko, L.; Sleziak, P.; Danko, M.; Bičárová, S.; Pociask-Karteczka, J. Analysis of changes in hydrological cycle of a pristine mountain catchment. 1. Water balance components and snow cover. J. Hydrol. Hydromech. 2020, 68, 180–191. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Catchment | Total Area [km2] | Total Forest Reduction [%] | C Study Area [ha] | ROC | DEB | WAT | ALM | DWP | IPS | REF | EXT | FIR | NEX | REX |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Color in Figure 1 |  | |||||||||||||
| MP | 83.0 | 5.0 | 758.7 | 11.6 | 16.6 | 0.8 | 25.7 | 21.2 | 2.1 | 14.6 | 4.6 | 0.0 | 0.0 | 0.01 |
| VP | 58.0 | 20.0 | 1031.7 | 13.3 | 10.0 | 0.3 | 14.1 | 16.8 | 7.3 | 8.1 | 27.5 | 0.0 | 1.7 | 0.01 |
| SP | 43.0 | 32.0 | 612.1 | 5.6 | 15.1 | 0.1 | 16.0 | 18.7 | 6.3 | 1.4 | 7.9 | 27.7 | 0.9 | 0.01 |
| Ecosystem Type | Abbreviation | Number of Points | Altitude (m a.s.l.) |
|---|---|---|---|
| Alpine meadow | ALM | 3 | 1730 |
| Dwarf pine | DWP | 5 | 1550 |
| Bark beetle attacked forest | IPS | 10 | 1180 |
| Mature undisturbed forest | REF | 20 | 1200 |
| Windthrow extracted | EXT | 24 | 1100 |
| Burnt windthrow | FIR | 8 | 1100 |
| Windthrow non-extracted | NEX | 8 | 1100 |
| 1-year-old windthrow | REX | 8 | 1100 |
| Code | Definition |
|---|---|
| MA3 | coefficient of variation in daily flows |
| MA44 | variability in annual flows divided by median annual flows; the variability is calculated as 90th minus the 10th percentile |
| ML13 | coefficient of variation in minimum monthly flows |
| ML22 | mean annual minimum flows divided by catchment area |
| MH17 | mean of the 25th percentile from the flow duration curve divided be median daily flow across all years |
| MH20 | mean annual maximum flow divided by catchment area |
| FL3 | total number of low flow spells (threshold equal to 5% of men daily flow) divided by the record length in years |
| FL2 | coefficient of variation in the low flow pulse count (below 25th percentile) |
| FH3 | high flow pulse count (high flow pulse is 7 times the median daily flow) |
| FH5 | mean of high flow events per year (high flow is 1 times the median flow) |
| DL6 | coefficient of variation in annual minima (1-day annual minima) |
| DL13 | mean annual 30 day minimum divided by median flow |
| DH12 | mean annual 7-day maximum divided by median flow |
| TL1 | mean Julian date of the 1-day annual minimum flow over all years |
| RA9 | coefficient of variation in number of reversals |
| RA8 | number of positive and negative changes in water conditions from one day to another (flow reversals) |
| Flux/ Ecosystem | ALM | DWP | IPS | REF | EXT | FIR | NEX | REX |
|---|---|---|---|---|---|---|---|---|
| GPP | −230 ± 8.2 | −886 ± 17.6 | −605 ± 19.4 | −1223 * | −1198 ± 24.5 | −1420 ± 33.0 | −1250 ± 29.1 | −425 ± 16.5 |
| ER | 220 ± 43.7 | 695 ± 90.3 | 1100 ± 175.0 | 1148 ± 120.1 | 846 ± 77.4 | 957 ± 175.0 | 909 ± 87.4 | 755 ± 96.4 |
| NEE | −10 ± 44.5 | −191 ± 91.9 | 495 ± 176.0 | −75 ± 120.1 | −352 ± 81.2 | −463 ± 178.1 | −341 ± 92.1 | 330.0 ± 97.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fleischer, P., Jr.; Holko, L.; Celer, S.; Čekovská, L.; Rozkošný, J.; Škoda, P.; Olejár, L.; Fleischer, P. Carbon Balance and Streamflow at a Small Catchment Scale 10 Years after the Severe Natural Disturbance in the Tatra Mts, Slovakia. Water 2020, 12, 2917. https://doi.org/10.3390/w12102917
Fleischer P Jr., Holko L, Celer S, Čekovská L, Rozkošný J, Škoda P, Olejár L, Fleischer P. Carbon Balance and Streamflow at a Small Catchment Scale 10 Years after the Severe Natural Disturbance in the Tatra Mts, Slovakia. Water. 2020; 12(10):2917. https://doi.org/10.3390/w12102917
Chicago/Turabian StyleFleischer, Peter, Jr., Ladislav Holko, Slavomír Celer, Lucia Čekovská, Jozef Rozkošný, Peter Škoda, Lukáš Olejár, and Peter Fleischer. 2020. "Carbon Balance and Streamflow at a Small Catchment Scale 10 Years after the Severe Natural Disturbance in the Tatra Mts, Slovakia" Water 12, no. 10: 2917. https://doi.org/10.3390/w12102917