A Fuzzy Inference System for Seagrass Distribution Modeling in the Mediterranean Sea: A First Approach



Laboratory of Ecological Engineering and Technology, Department of Environmental Engineering, Democritus University of Thrace, 67100 Xanthi, Greece
*
Author to whom correspondence should be addressed.
Water 2020, 12(10), 2949; https://doi.org/10.3390/w12102949
Submission received: 18 August 2020
/
Revised: 15 October 2020
/
Accepted: 17 October 2020
/
Published: 21 October 2020
(This article belongs to the Special Issue Observations and Models for End-User Services in Coastal Marine Systems)
Abstract
:A Mamdani-type fuzzy-logic model was developed to link Mediterranean seagrass presence to the prevailing environmental conditions. UNEP-WCMC (seagrass presence), CMEMS, and EMODnet (oceanographic/environmental) datasets, along with human-impact parameters were utilized for this expert system. The model structure and input parameters were tested according to their capacity to accurately predict the presence of seagrass families at specific locations. The optimum Fuzzy Inference System (FIS) comprised four input variables: water depth, sea surface temperature, nitrates, and bottom chlorophyll-a concentration, exhibiting reasonable precision (76%). Results illustrated that Posidoniaceae prefers cooler water (16–18 °C) with low chlorophyll-a levels (<0.2 mg/m3); Zosteraceae favors similarly cooler (16–18 °C) and mesotrophic waters (Chl-a > 0.2 mg/m3), but also slightly warmer (18–19.5 °C) with lower Chl-a levels (<0.2 mg/m3); Cymodoceaceae lives in warm, oligotrophic (19.5–21.0 °C, Chl-a < 0.3 mg/m3) to moderately warm mesotrophic sites (18–21.3 °C, 0.3–0.4 mg/m3 Chl-a). Finally, Hydrocharitaceae thrives in the warm Mediterranean waters (21–23 °C) of low chlorophyll-a content (<0.25 mg/m3). Climate change scenarios show that Posidoniaceae and Zosteraceae tolerate bathymetric changes, and Posidoniaceae and Zosteraceae are mostly affected by sea temperature rise, while Hydrocharitaceae exhibits tolerance at higher sea temperatures. This FIS could aid the protection of vulnerable seagrass ecosystems by national and regional policy-makers and public authorities.
1. Introduction
Seagrasses are the only submerged marine plants with an underground root and rhizome system forming beds and meadows. They play a key role in the ecosystem services of the global coastal zone in relation to nutrient biogeochemical cycling, carbon sequestration, sediment stabilization, fish refugia, and food-web structure [1]. Although the global species diversity of seagrasses is relatively low (<60 species), understanding their distribution is particularly important for ecologists developing bioregional models, covering all oceans and climatic zones together with the respective species assemblages [2]. Most aquatic ecosystem health assessment studies rely on seagrass species richness and distribution, since these serve as valuable bio-indicators reflecting recent environmental changes, especially the release of pollutants and eutrophication events. Halophila minor and Halophila ovalis act as bio-indicators for trace metal pollution and sediment accumulation [3], and Zostera marina is an eutrophication indicator [4], while the genus Cymodocea acts as a heavy-metal bioaccumulator and tolerant bioindicator of pollution [5], rapid coastal development, and human intervention [6]. Finally, Posidonia oceanica meadows are directly linked to the degree of human impact, like effluents, many nutrients, organic matter, trace metals, coastal settlements, fish farms, and trawling [7,8,9].
It is evident that seagrass species face significant challenges due to their high vulnerability related to a variety of anthropogenic disturbances concentrated along the coastal zone [2], and their sensitivity to environmental changes driven by climate change [10], ultimately leading to their global decline [11]. In the Mediterranean bioregion, there are eight seagrass species, namely Cymodocea nodosa, Halophila decipiens, Halophila stipulacea, Posidonia oceanica, Ruppia cirrhosa, Ruppia maritima, Zostera noltii, and Zostera marina, with four of six families declining in the western Mediterranean region, implying that priority conservation policies are needed [11,12]. To understand the complex interrelation between oceanographic, environmental, morphodynamic, and human impact conditions and the distribution of seagrass species, modern data-driven models have been developed and implemented, following Machine Learning and Artificial Neural Networks techniques [13,14]. Such models are capable of exploring the seagrass presence/absence dynamics, and link seagrass families and species to the main environmental drivers determining their distribution, and ultimately, their abundance.
The exploitation of patterns between seabed habitat types and environmental parameters is particularly important for ecologists, since the abundance and distribution of seabed biological communities are directly linked to water column dynamics and quality [15]. In parallel, seagrass presence/absence and internal body growth asymmetries provide valuable metrics for the evaluation of marine ecosystems’ ecological status [16]. Although the interrelation between physical, chemical, biological, seabed- and human-related drivers to seagrass species distribution is strongly non-linear, and therefore difficult to identify, modern data-driven models and techniques explore these hidden patterns. In addition, the development of systematic, freely available, diverse databases over the latest decades, and the development of tools for Big Data fusion and aggregation, lead to the better understanding of coastal benthic processes and human impacts.
In the present study, fuzzy logic modeling explored the nonlinear dynamics between the environment-ecosystem-human gradient and its impact on seagrass distribution in the Mediterranean Sea. A Fuzzy Inference System (FIS) is the process of formulating the mapping between a given set of inputs to an output using fuzzy logic. Such systems are particularly suited to model the relationship between variables in environments that are either ill-defined or very complex, as the case of seagrass distribution and its relation to environmental and human drivers. The Fuzzy Inference System (FIS) is an engine that applies reasoning to compute fuzzy outputs, and involves a knowledge base which defines rules and membership functions (MFs). The system is built using a set of “if–then” rules, which, like any conventional rule in artificial intelligence, has the general form, “If x is A, then z is C”, where A and C are linguistic values defined by fuzzy sets in the universes of discourse X and Z, respectively. The “if” and “then” parts are called the antecedent and the consequent of a rule, respectively.
The optimal model generated was further used to assess the response of the main Mediterranean seagrass species distribution patterns to changes in climate (gradual water temperature increase, sea level rise) and eutrophication. This FIS model could be used by policy-makers and public authorities responsible for the Marine Strategy Framework Directive (MSFD) implementation to understand and assess pressures and impacts on seagrass ecosystems, link abiotic factors on determining the appropriate Good Environmental Status (GES) and establishing baselines and targets for biodiversity assessments in the frame of MSFD (Descriptor 1) and UN Sustainability Development Goals (Goal 14.2), in line with the prevailing hydrologic, oceanographic, biogeographic, and climatic conditions.
The work was conducted in the framework of the Horizon 2020 ODYSSEA Project. Some of the aims of this project include developing appropriate algorithms, aggregating data from diverse databases, revealing hidden relationships among abiotic and biological parameters, and identifying species distribution patterns and species richness trends.
2. Materials and Methods
2.1. The Seagrass Dataset
The initial dataset was the Global Distribution of Seagrasses dataset, produced by the United Nations Environment Programme—World Conservation Monitoring Centre (UNEP-WCMC) [17,18]. The dataset illustrates the global distribution of 73 seagrass species and has the form of a geo-referenced shapefile composed of two subsets of points and polygons, indicating the occurrences measured from 1934 to 2015 on seagrass family, genus, and species. A total of 1771 locations were found for the major Mediterranean seagrass families (Cymodoceaceae, Zosteraceae, Posidoniaceae, Hydrocharitaceae, Ruppiaceae) (Table 1).
A limited number of problematic points were identified, based mostly on the depth zone distribution and the distance to the coast. Data were filtered on Mediterranean Sea seagrass families, and ultimately, the species. Ruppiaceae data were excluded from analysis due to their limited occurrence (<2.0%).
2.2. The Environmental Drivers Dataset
The distribution of seagrasses in the Mediterranean Sea is assumed to vary in relation to a series of physical, chemical, biological, and seabed- and human-related parameters. For each geolocation in which seagrass species are reported, hydrographic data (water temperature, salinity, currents, waves) and water quality data (nutrients, dissolved oxygen, chlorophyll-a, net primary production rates) at the surface and bottom of the water column were retrieved from the Copernicus Marine Environmental Service (CMEMS, https://marine.copernicus.eu/) data products. These are gridded, mean-monthly oceanographic data, covering the whole Mediterranean Sea during variable periods from 1987 to 2015 (Table 2). The only exception was the wave dataset that was reported on an hourly basis and subsequently converted into mean-monthly values.
Other factors potentially affecting the distribution of seagrass species are the water depth and the substrate conditions (varying from mud to rock) which were provided by the European Marine Observation and Data Network (EMODnet, https://www.emodnet.eu/en).
Finally, the distance of each seagrass observation point to the closest point of human influence (port, coastal city, river mouth, distance to coast) was computed, using the haversine distance method.
The final dataset consisted of 573 data points, representing each location (geographical coordinates) with the presence of the four main Mediterranean seagrass families, namely Zosteraceae, Hydrocharitacheae, Cymodoceaceae, and Posidoniaceae, together with the climatological (multi-year) means produced by averaging the mean-monthly values of physical, water quality, and biological parameters [19]. Bathymetry, substrate properties, and distances of seagrass locations from areas of potential human impact (coast, cities with a population of over 50,000 inhabitants, ports, and river mouths) were also included in the dataset. These parameters are considered as potential drivers to explain marine flora distribution and the response to environmental changes. The dataset was randomly divided into two parts: 90% was defined as a training dataset, while the remaining 10% acted as a validation dataset.
2.3. The Fuzzy Inference System
A Mamdani-type fuzzy inference system was developed for this study, which comprised four steps (Figure 1): fuzzification of input variables, rules construction and evaluation, aggregation of rules output, and defuzzification [20,21].
The FIS consisted of four physical, chemical, biological, seabed- and human-related input parameters, a series of well-designed fuzzy rules, based on the most frequent parameters interrelated in the training set, and receiving one aggregate value representing the seagrass family abundance. The FIS should be able to respond to the following question: “Given a series of environmental driving conditions prevailing in an area, what is the seagrass family favored by these conditions?”
The first step involves taking the crisp inputs, i.e., each numerical value, and assigning the appropriate fuzzy sets (defined here as Very Low, Low, Medium, High, and Very High) through the selected membership function (in this case, a trapezoidal membership function). The trapezoidal membership function used to transform crisp inputs into membership values belonging to the various fuzzy sets of the FIS has the form:
where a, b, c, and d are the membership function parameters (Table 3). Examples for bottom chlorophyll-a and surface temperature are shown in Figure 2 and Figure 3.
After fuzzification, the second step involves taking the fuzzified inputs and applying them to the antecedents in a series of constructed fuzzy rules. If a given fuzzy rule has multiple antecedents, then the fuzzy operator (in this case, AND) is used to obtain a single number that represents the result of the antecedent evaluation. This number (the true value) is then applied to the consequent membership function. The IF-THEN fuzzy rules constructed according to the most frequent interrelation of input parameters observed in the training set have the following form. Rule 1: “If the water depth is Low, AND the water temperature at the sea bottom is Low, AND the chlorophyll-a concentration at the sea bottom is Low, AND the nitrates at the sea surface are Very Low, THEN the Seagrass Family favored in these conditions is Posidoniaceae’’.
A set of similar fuzzy rules may be developed and inserted in the FIS. When feeding the FIS with more data, that is, developing and adding more rules, the model provides more accurate estimates of the favored Seagrass Family presence. In our case, 45 total rules were used to associate the most frequently appearing independent antecedents in the training sets with a rule.
The third step involves the aggregation, a process that produces an overall output by considering the membership functions of all rule consequents and combining them into a single fuzzy set.
Finally, defuzzification is the process which leads to a final output as a crisp number. The input in the defuzzification process is the aggregate fuzzy set, and the output is a single number. There are several defuzzification methods, but the most popular one is the centroid technique [20,21], which is followed here. The seagrass family output is produced through a triangular membership function, of the form:
The defuzzification membership functions of the FIS are discrete, resulting in a definite seagrass species. If the output falls in the interval [0, 0.25], then the seagrass family is Zosteraceae; if [0.25, 0.50], then the seagrass family is Hydrocharitaceae; if [0.50, 0.75], then it is Cymodoceaceae; and finally, in the range [0.75, 1.00], it is Posidoniaceae (Figure 4).
Several experimental Fuzzy Inference Systems were developed in this study, comprising different combinations of the four input parameters and keeping the one output parameter, that is, the seagrass family, constant. The FIS was developed using the Fuzzy Logic Toolbox operating under MATLAB 7.0 (The Mathworks, Inc., Natick, MA, USA).
2.4. Evaluation Metrics
The most commonly used measure to evaluate this type of FIS model is the Classification Precision of the system, being the fraction of relevant/correctly classified instances, in terms of seagrass family presence, over the validation dataset (10% or 58 records from the dataset).
Precision is a good metric for balanced datasets, like the case studied here for seagrass family presence [18], and is defined as:
where TP is known as the number of true (correct) assessments, and FP indicates the number of incorrect assessments produced by the examined FIS.
Therefore, the model returns the Mediterranean seagrass family that best suits the input environmental parameters. When the forecasted family coincides with values in the UNEP dataset, this instance is considered a true-positive. In case of an incorrect assessment, a false-positive instance is considered. For example, in Rovinj, Croatia, the FIS correctly assessed that the environmental drivers favor Zosteraceae, but in a location at the Gulf of Gabes, Tunisia, the assessment was incorrect (Zosteraceae instead of Cymodoceaceae).
2.5. Statistics
Aiming to detect possible statistical differences between the environmental conditions prevailing in sites with different seagrass families, a two-way ANOVA was performed. In the case of a significant difference between levels (p < 0.05), a Tukey’s post-hoc test was used to show which families differed.
2.6. FIS Sensitivity Analysis and Climate Change Scenarios
Sensitivity tests using the optimum FIS were conducted to simulate the impact of climate change and increased human pressure on seagrass communities under different scenarios, thus examining the level of seagrass tolerance and their progressive species replacement. Model runs incremented each parameter by −15%, −10%, −5%, +5%, +10%, and +15% of its initial value. Then, the optimum FIS was used to assess and predict the most appropriate seagrass family favoring these environmental conditions. The developed tests were “closed”, that is, seagrass families may change among them, but their disappearance is not allowed. This is a deficiency of the presently developed model that will be resolved in a future study.
3. Results
3.1. Data Analysis
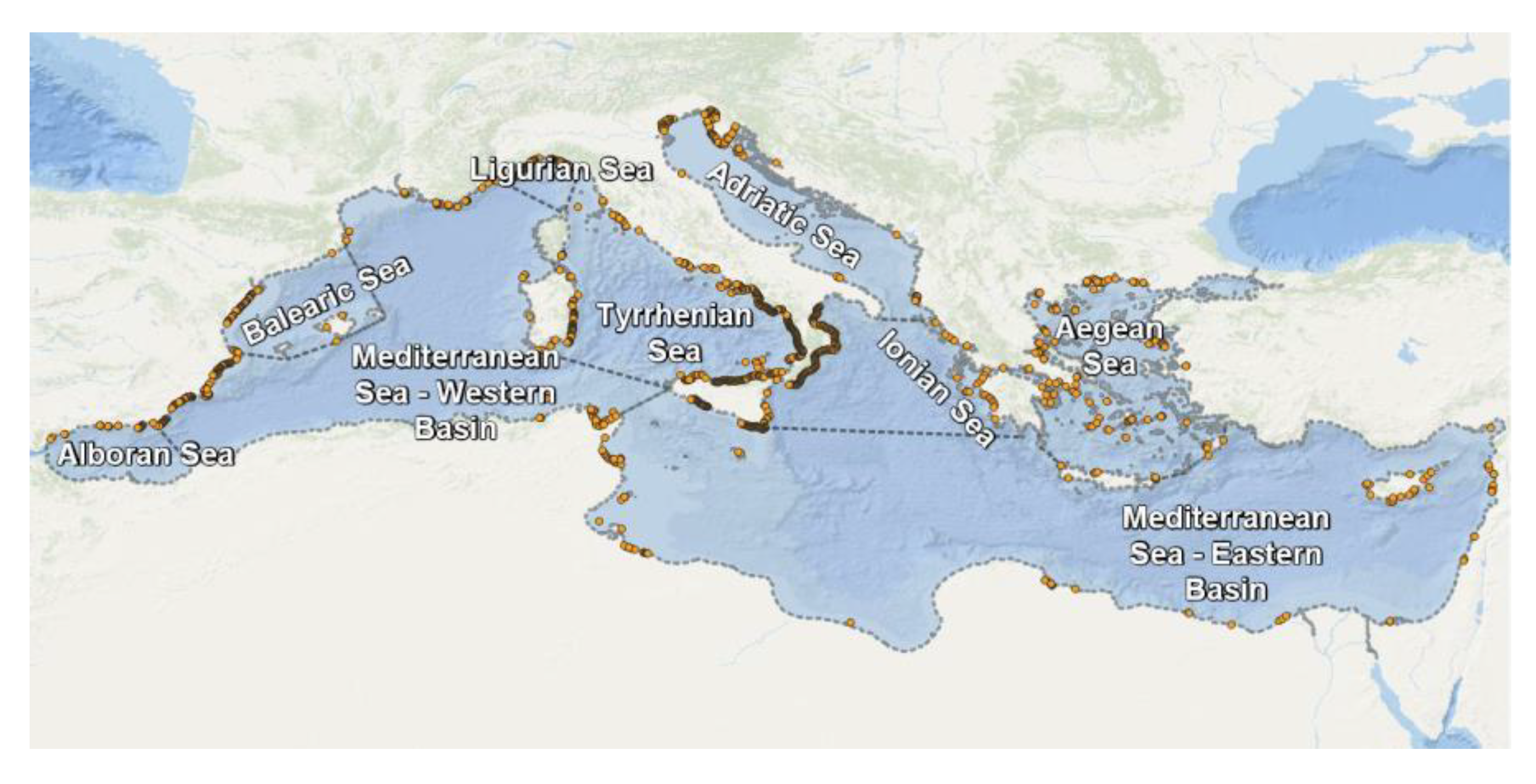
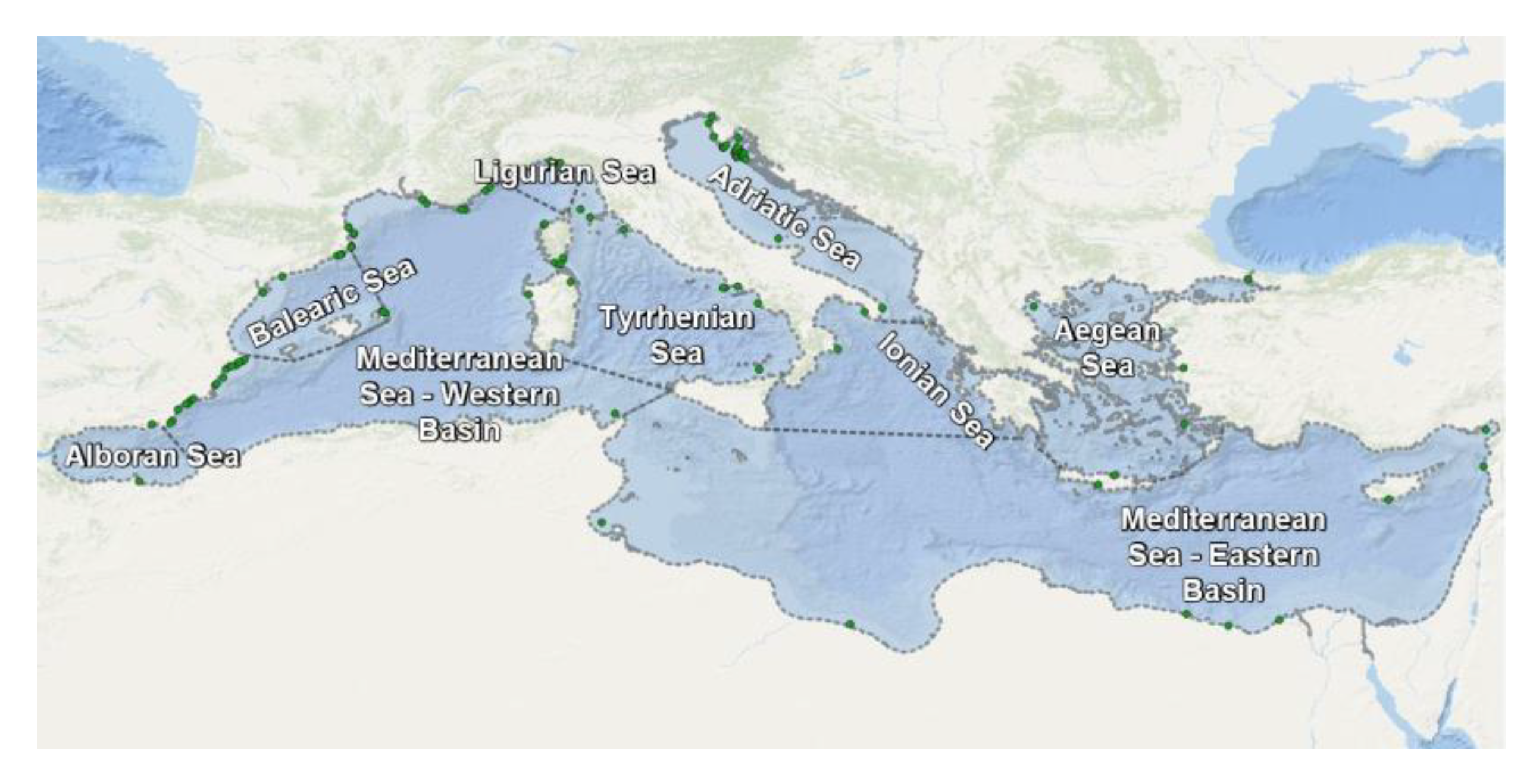
Seagrass geolocation data were imported and mapped using a QGIS (QGIS 3.14 Pi). Figure 5 and Figure 6 illustrate the spatial distribution of Cymodoceaceae and Zosteraceae in the Mediterranean Sea, respectively.
Overall, 34% of the Cymodoceaceae records are found in the Tyrrhenian Sea, 14% in the Western Mediterranean Basin, and 13% in the Adriatic Sea. The Posidoniaceae sites are located in the Adriatic Sea (38%) and 30% in the Western Mediterranean Basin, while the Zosteraceae were primarily in the Western basin (42%), the Adriatic (22%), and in the Tyrrhenian Sea (20%). Finally, all Hydrocharitaceae are found in the Eastern Mediterranean Basin.
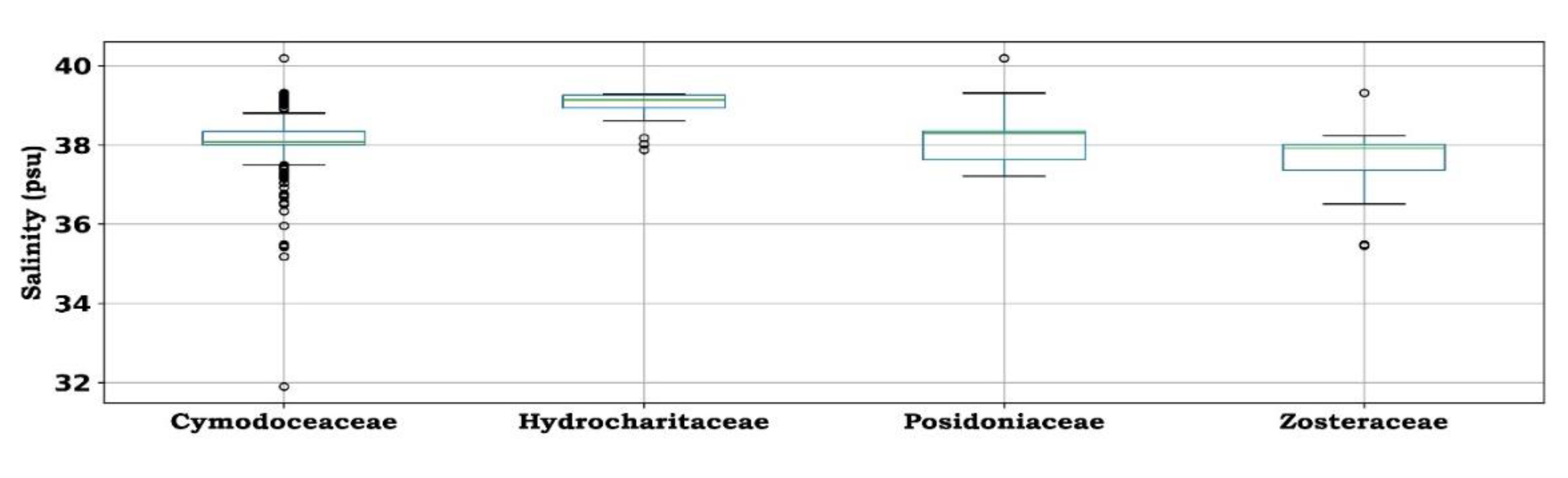
Hydrocharitaceae, and specifically the seagrass species Halophilla, favor the most saline environments, within the narrow yearly-averaged salinity range from 39 to 39.5 psu. Posidoniaceae and Cymodoceaceae prefer similar salinity levels, ranging between 37.3 and 39.3 psu. The interquartile Hydrocharitaceae and Cymodoceaceae range exhibits a “tighter” distribution, while Posidoniaceae, Ruppiaceae, and Zosteraceae favor marine environments within a wider salinity range (Figure 7).
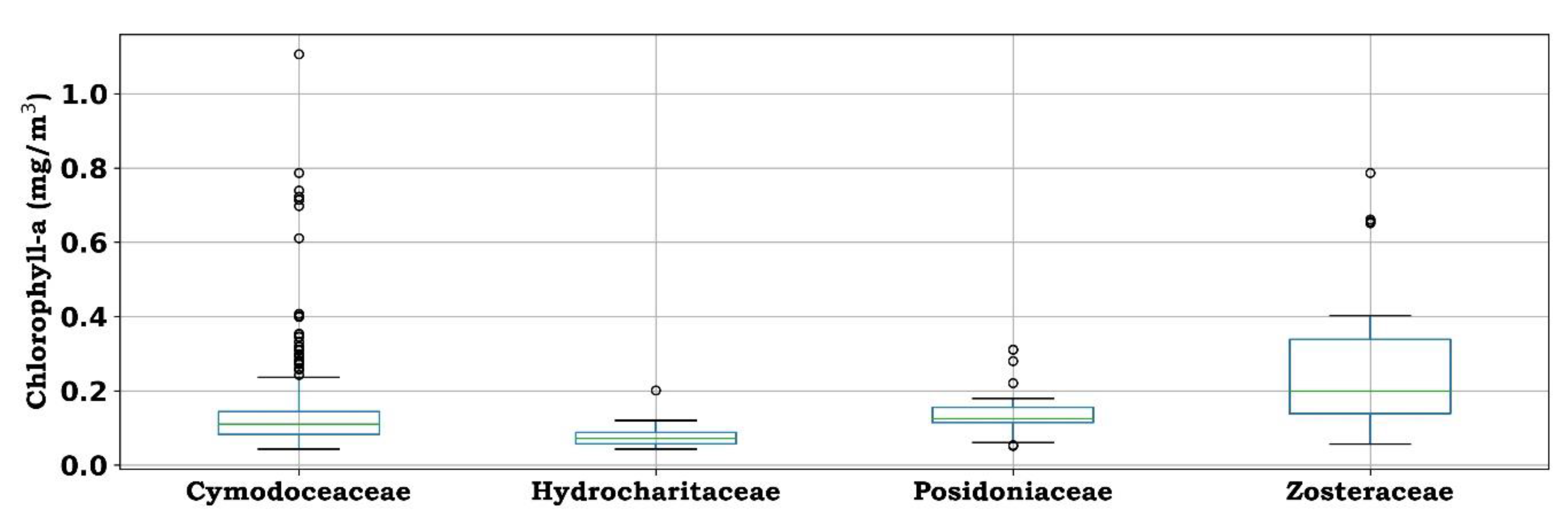
Similarly, each family prefers different ranges of chlorophyll-a concentration. For Hydrocharitaceae and Posidoniaceae, they are typically found within a narrower range of chlorophyll-a levels (0.07 ± 0.02 mg/m3 and 0.13 ± 0.05 mg/m3, respectively), while other families occur in habitats with a wider range of chlorophyll-a levels (e.g., Zosteraceae: 0.26 ± 0.16 mg/m3), explaining their tolerance to eutrophication (Figure 8).
3.2. Experimental Tests Analysis
In the initial experimental test, four input parameters were imported into the developed FIS: the distance from major cities, the concentration of phosphates at sea surface, the seabed salinity, and the benthic concentration of chlorophyll-a. Some basic fuzzy rules of this initial test are shown in Table 4. The interpretation of the first rule is: “If the Distance from Cities is Medium, the concentration of Surface Phosphates is Very Low, the level of Bottom Salinity is Medium, and the concentration of Bottom Chlorophyll-a is Low, then Zosteraceae is favored”. This experimental test showed limited precision (52%). After rule improvements and excluding Posidoniaceae from the model, due to the limited number of records in the training dataset, model precision reached 63% in the testing/validation dataset.
In another test, the FIS was constructed from the distance of each seagrass location from the coastline, bottom water temperature, salinity, and the benthic chlorophyll-a concentration. Despite improvements in overlapping fuzzy rules, this model reached precision of only 61% in the validation procedure. Overall, 22 experimental FIS tests were developed and examined for their capacity in predicting the environmental conditions preferred by the different seagrass families.
The optimum FIS included water depth, sea surface temperature, surface nitrate concentration, and bottom chlorophyll-a concentration (Table 5). This FIS exhibited precision of 72%, and after fuzzy rules’ manipulation and adjustment, like omitting some fuzzy rules of quite similar content, the FIS reached a precision level of 76%. However, model precision was not evenly distributed over the Mediterranean. Higher precision (82%) was found in the eastern Mediterranean Sea, especially along the coastal zones of the Adriatic, Ionian, and Aegean Seas. Model precision decreased to 68% in the western basin, mostly in the Alboran and Tyrrhenian Seas, as well as the coastline of Northern Africa.
3.3. Seagrass Family-Preferred Environments
Merging the seagrass database of seagrass occurrence within the Mediterranean with the CMEMS and EMODnet data revealed the most favorable environmental conditions for each seagrass family using the optimum FIS model. Posidoniaceae, driven mostly by P. oceanica, prefers cooler waters from 16–18 °C and a low chlorophyll-a presence (<0.2 mg/m3). Zosteraceae seabeds also favor cooler (16–18 °C) temperatures in more mesotrophic waters (Chl-a > 0.2 mg/m3), but are also found in slightly warmer waters (18–19.5 °C) with lower Chl-a levels (<0.2 mg/m3). Cymodoceaceae are found over a wide range of environmental conditions, from warm, oligotrophic waters (19.5–21.0 °C and Chl-a < 0.3 mg/m3) to moderately warm mesotrophic areas (18–21.3 °C and 0.3–0.4 mg/m3 Chl-a). Finally, Hydrocharitaceae seem to thrive in warmer Mediterranean waters (mean annual temperature 21–23 °C) with lower chlorophyll-a concentrations (<0.25 mg/m3) (Figure 9).
3.4. Results from Climate Change Scenarios
Scenario 1 examined the impacts produced by changing the bathymetry in the locations with seagrass beds. The most sensitive seagrass families were Hydrocharitaceae and Cymodoceaceae. The reduction in water depth by 10% and 15% led to the reduction in the relative abundance of Hydrocharitaceae by 12% and 14%, respectively, being replaced by Cymodoceaceae. In contrast, sea level rise seemed to have little effect on all seagrass families.
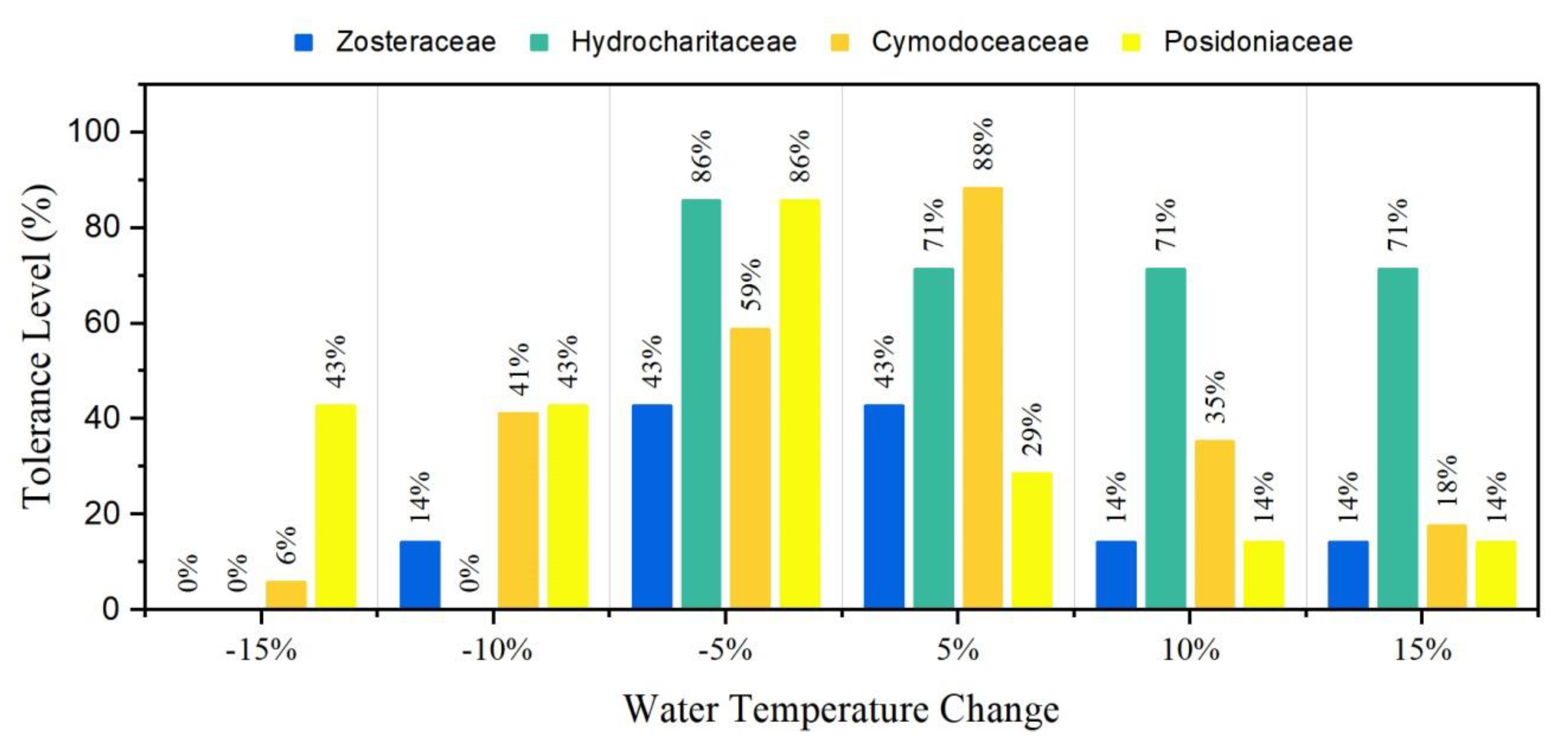
Scenario 2 studied the response of seagrass families to water temperature changes. All four families were sensitive to these variations. Posidoniaceae exhibited higher tolerance to decreasing mean water temperature, while Hydrocharitaceae had higher tolerance to sea temperature rise (Figure 10). More specifically, 43% and 57% of the locations with Posidoniaceae were replaced by Cymodoceaceae when the water temperature rose by 10% and 15%, respectively. In parallel, 35% of the Cymodoceaceae sea beds were replaced by Posidoniaceae as water temperature decreased by 15%. Another 18% of these Cymodoceaceae meadows changed into Zosteraceae under these conditions. Posidoniaceae replaced 43% of the initial Zosteraceae seabeds when local water temperature dropped by 15%. In contrast, as water temperature rose by 5%, Zosteraceae seabeds changed into Hydrocharitaceae and Cymodoceaceae by 43% and 14%, respectively.
Some seagrass meadows at family level in the Mediterranean were tolerant to water temperature change. Posidoniaceae is more tolerant in cooler water temperature environments, while Hydrocharitaceae and Cymodoceaceae meadows remain relatively stable under mild sea temperature increases, allowing only the former to sustain the higher water temperature increase.
The final scenario examined the response of seagrasses to changes in bottom chlorophyll-a levels. Hydrocharitaceae remained unaffected by changes in chlorophyll-a levels. Lower levels in chlorophyll-a led to the replacement of Zosteraceae by Hydrocharitaceae (29% and 40%, with a 5% and 10% reduction, respectively) and Cymodoceaceae (29% under 15% drop). On the other hand, an increase of 15% in the chlorophyll-a annual levels led to a reduction of 14% of the Posidoniaceae meadows.
4. Discussion
In this work, a fuzzy logic model was developed using existing datasets for the Mediterranean basin to examine the main environmental parameters affecting the distribution of seagrass families. Fuzzy logic models of the Mamdani type have never been implemented for such a complex task. In a similar study, [13] considered a series of morphodynamic, environmental, and human impact variables while employing Machine Learning algorithms to predict the presence–absence of P. oceanica seagrass species. Their dataset, however, was limited and rather unbalanced, biasing absence records and affecting the model’s reliability. Results showed that P. oceanica presence/absence patterns depended on nitrates and silicates, water depth, mean sea surface temperature, salinity, and distance to river mouths. Although chlorophyll-a concentrations were not included in the analysis, the correctly classified instances reached 78.3% using the random forest classifier.
A more comprehensive study was performed by [14,19] detecting seagrass presence/absence and distinguishing seagrass families in the Mediterranean through supervised learning methods. In these papers, chlorophyll-a and distance to the coast appeared more relevant in explaining seagrass presence/absence, while chlorophyll-a, salinity, distance to major cities, and nutrients were found as the main drivers for detecting the presence of different seagrass families.
The improvement of the present model can be explained by (a) the simplicity of using only four imported environmental predictors compared to 217 in [14], thus achieving higher precision when minimizing the impact of irrelevant variables; (b) predictors were aggregated to their long-term means, compared to the use of mean-monthly variables representing limited time-intervals (e.g., in [14] Chl-a for just December 2015 was used); and (c) the FIS does not operate like a “Black-box” ML model in which the lack of transparency and accountability may lead to poor interpretability. In fuzzy systems, rules are user-constructed according to data reasoning, they are flexible, and can be modified or receive different weights, making the model easier to interpret. The [14,19] model tested several machine-learning classifiers and achieved the highest precision of 44.4% on seagrass family classification with the random forest algorithm. The present FIS, although simpler, achieved a precision level of 76%.
This model consisted of four main drivers: water depth, bottom chlorophyll-a concentration, surface water temperature, and nitrates. Although [14,19] used a different environmental driver dataset, quite similar predictors were found (bathymetry and chlorophyll-a). Moreover, in this work, water temperature was found to play a major role in determining seagrass family presence, instead of salinity in [14]. The fact that long-term climatological means were used as predictors in this work, compared to mean-monthly values imported in the machine learning model of [14,19] seems responsible for this relative weight change among variables. In the Mediterranean, the variability in water temperature is more important than salinity variability among the various seagrass locations, and this was captured by the FIS. The influence of water temperature on P. oceanica growth was also highlighted in [22], with limited presence of this species in the warmer Levant Sea. The impact of nutrients was found to be important in all similar works [13,14,19], since food availability determines seagrass growth, distribution, and metabolism [23].
Water depth is an important parameter to predict the location of seagrass families, since it indirectly expresses changes in seabed temperature, pressure, light availability, and wave disturbance [24]. Of the above-listed factors, seabed light availability appears to be the most influential driver in determining the colonization depth of various seagrass species [25], since it affects the photosynthetic activity. In the Mediterranean, Cymodoceaceae, mostly C. nodosa, was found over a broad range of depths, from shallow waters to depths of 60 m in sheltered to semi-exposed coasts, while P. oceanica was also present down to 50 m [26]. In the present dataset, Cymodoceaceae and Zosteraceae were found to be a shallow water species (mean depth 17 m), while Posidoniaceae was found in moderate depths (mean ± sd: 32 ± 16 m), and Hydrocharitaceae in moderate to deep water (66 ± 28 m). Tukey’s post hoc test (p < 0.05) showed that Posidoniaceae sites were found to be significantly different in terms of bathymetry to Zosteraceae and Cymodoceaceae habitats. Above-substrate biomass, leaf biomass, and shoot density of both species was found to decline from shallow to higher depths in the Spanish Mediterranean Sea [27], probably due to changes in light intensity and seabed temperature, explaining that bathymetry is an important factor capable of distinguishing seagrass families, as shown by the present FIS.
Water temperature is also a significant driver for seagrass distribution, influencing growth rates, reproductive patterns, and enzymic and metabolic functions when found within a physiological optimum. Extreme heat may enhance mortality for a number of seagrass families [28]. The optimum FIS clarified sea water temperature preferences by family in the Mediterranean: Posidoniaceae and Zosteraceae favor cooler temperatures (16–18 °C), Cymodoceaceae are found in moderate (19.5–21.0 °C) to warm waters (18–21.3 °C), while Hydrocharitaceae grow in warmer areas (21–23 °C). Statistical tests (Tukey’s post hoc test, p < 0.05) illustrated that all seagrass habitats were characterized as significantly different in terms of water temperature, except for Zosteraceae and Posidoniaceae. These findings are in agreement with the species distribution patterns over the Mediterranean Sea, with warm-water species mostly being along the North African coastline and cold-water species being along the northern shores of the Aegean, Ionian, Adriatic, and Tyrrhenian Seas [13]. Temperature preferences may determine which species will be affected under rising temperature climate scenarios. Under such conditions, the decline of one species may lead to the replacement by another [29], following the patterns defined by the FIS. In some examples of disturbance, no recovery was observed, although this case was not examined by the FIS.
Chlorophyll-a levels were important for seagrass family distribution. Each family prefers specific and different values of chlorophyll-a (Figure 8). Posidoniaceae and Hydrocharitaceae favor oligotrophic systems; Zosteraceae is abundant in mesotrophic environments; while Cymodoceaceae is tolerant to a wide range of chlorophyll-a concentrations. According to [14], chlorophyll-a levels in winter months (mostly in December) is the key parameter determining seagrass presence/absence and family identification. According to [22], P. oceanica meadows are absent in the vicinity of river mouths due to increased turbidity and chlorophyll-a levels. During eutrophication events, seagrass suffers from reduced light conditions, increased epiphyte growth, decline in dissolved oxygen, and anaerobic organic matter decomposition, leading ultimately to sulfide stress that few species can tolerate [30]. P. oceanica is very sensitive to sulfide stress [31], whereas temperate species growing on terrigenic sediments, such as C. nodosa and Z. marina, showed only a minor response to this stress [32].
Nutrients, mostly in the form of nitrates, also represent a key environmental driver for seagrass species abundance, since food availability controls seagrass growth, distribution, and metabolism [33]. Several nutrient sources, mostly rivers, outflow along the Mediterranean coastline, providing the appropriate nutrient levels to seagrass sustainability. In eutrophic waters, phytoplankton is favored at the expense of seagrass production, resulting in the loss of seagrass cover at high nutrient concentrations [32]. The effect of seagrasses on nitrogen cycling and uptake kinetics is species-dependent, where some species exhibits increased, and others reduced uptake of affinities and rates [32]. This supports the finding that nitrogen concentrations may act as an environmental driver for seagrass species differentiation, as produced by the optimum FIS. Nitrogen assimilation by seagrasses requires the nitrate reduction to ammonium, a process taking place with higher rates in leaves than roots. This allows the easier utilization of dissolved inorganic ammonium as the “preferred” pool of nutrients, compared to the ammonium pools in sediments and pore water [32]. Morphological leaf variations among seagrass families, like leaf width, length, number of leaves per shoot, and so forth support the variable dissolved nitrate and ammonium uptake rates from the benthic waters, in agreement with the selection of the FIS. The main difference from study [14] is that the present FIS identified nitrates as an important factor compared to the machine-learning model that emphasized phosphates. Seagrass beds are considered significant phosphorus sinks, with variable, species-dependent rates of uptake and assimilation.
In terms of seagrass resilience to climate change impacts, the optimum FIS indicated that: (a) Zosteraceae is highly tolerant to changes in water depth and nutrient levels, as well as to small changes in chlorophyll-a levels, but extremely sensitive to temperature changes; (b) Hydrocharitaceae is resistant to increases in water temperature, chlorophyll-a, and nutrient levels, but sensitive to bathymetric changes; (c) Cymodoceaceae meadows remain relatively stable under mild sea temperature increases, replacing Posidoniaceae under strong sea water temperature rises; and (d) Posidoniaceae appears threatened by an increase in water temperature, favors oligotrophic waters, and is being found in moderate water depths.
Since the proposed model (in terms of input variables) follows the “trial and error” approach, it could be further improved through extended testing. The inclusion of taxa disappearance, as an additional output state, is also another option, advancing the FIS and making the model more realistic. However, expert knowledge is needed for such advancement. Other tests could include finding the most accurate shape and boundaries of the membership functions and examining the most appropriate defuzzification method.
5. Conclusions
This work has developed a simple but novel self-learning expert-system application, based on Mamdani fuzzy logic to predict the occurrence of Mediterranean seagrass habitats at the family level, according to the environmental conditions prevailing in an area. The system was developed utilizing diverse databases, from UNEP-WCMC for seagrass distribution and CMEMS and EMODnet for environmental conditions at each seagrass site. The optimum model receives input values from four parameters, namely water depth, sea surface temperature, surface nitrates, and bottom chlorophyll-a, and identifies seagrass family distribution patterns with fair classification precision (~76%).
The present FIS is capable of describing favorable living conditions for seagrass families across the Mediterranean Sea. Posidoniaceae, and mostly P. oceanica prefers cool, oligotrophic waters; Zosteraceae seabeds favor a wider temperature range (16–19.5 °C); and mesotrophic waters, Cymodoceaceae, and mostly C. nodosa appear tolerant to a broad range of living conditions, from warm, oligotrophic, to moderately warm mesotrophic areas; while Hydrocharitaceae prefer the warmer, oligotrophic parts of the Mediterranaean.
The FIS also has the capability to identify the impact of environmental change on seagrass habitats as those induced by climate change, illustrating that (a) Hydrocharitaceae and Cymodoceaceae are the most sensitive families to increases in water depth with sea level rise, while Posidoniaceae and Zosteraceae are tolerant to the climate change drivers examined; (b) Cymodoceaceae is the family with higher tolerance to mild (+5%) increases in sea temperature; (c) Hydrocharitaceae exhibits tolerance to higher (+10–15%) increases in sea temperature; (d) Posidoniaceae and Zosteraceae are mostly affected by temperature rise at any level, and (e) Posidoniaceae exhibits higher tolerance to a decrease in mean water temperature.
Based on these model results, it is evident that tolerance toward disturbances, as well as growth and recolonization potentials differ among various seagrass species, and distribution patterns can now be predicted based on these taxonomic tolerances. Seagrass presence requires several environmental conditions to be satisfied, and quantifying the relative importance of these conditions is the primary challenge for predicting seagrass habitat suitability. Thus, the present FIS model could be potentially used by policy-makers in order to (a) aid the design of a conservation action plan for areas that seagrass species are present; (b) explore the environmental factors and improve those needed in areas where seagrasses are absent, thus supporting ecosystem restoration; and (c) establish baselines and targets for biodiversity assessments required for MSFD implementation.
Author Contributions
Conceptualization, G.S.; methodology, G.S.; software, G.S.; validation, D.P., N.K., G.S; formal analysis, D.P.; resources, N.K.; data curation, N.K.; writing—original draft preparation, G.S.; writing—review and editing, N.K., G.S.; visualization, D.P.; supervision, G.S.; project administration, G.S.; funding acquisition, G.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by European Union’s Horizon 2020 Research and Innovation Program (H2020-BG-12-2016-2), grant number No. 727277—ODYSSEA (Towards an integrated Mediterranean Sea Observing System). The article reflects only authors’ view and that the Commission is not responsible for any use that may be made of the information it contains.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Barnes, R.S.K.; Claassens, L. Do beds of subtidal estuarine seagrass constitute a refuge for macrobenthic biodiversity threatened intertidally? Biodivers Conserv. 2020, in press. [Google Scholar] [CrossRef]
- Short, F.; Carruthers, T.; Dennison, W.; Waycott, M. Global seagrass distribution and diversity: A bioregional model. J. Exp. Mar. Biol. Ecol. 2007, 350, 3–20. [Google Scholar] [CrossRef]
- Ahmad, F.; Azman, S.; Said, M.I.M. Lavania-Baloo Tropical seagrass as a bioindicator of metal accumulation. Sains Malays 2015, 44, 203–210. [Google Scholar] [CrossRef]
- Lee, K.-S.; Short, F.T.; Burdick, D.M. Development of a nutrient pollution indicator using the seagrass, Zostera marina, along nutrient gradients in three New England estuaries. Aquat. Bot. 2004, 78, 197–216. [Google Scholar] [CrossRef]
- Ferrat, L.; Pergent-Martini, C.; Roméo, M. Assessment of the use of biomarkers in aquatic plants for the evaluation of environmental quality: Application to seagrasses. Aquat. Toxic. 2003, 65, 187–204. [Google Scholar] [CrossRef]
- Orfanidis, S.; Papathanasiou, V.; Gounaris, S.; Theodosiou, T. Size distribution approaches for monitoring and conservation of coastal Cymodocea habitats. Aquat. Conserv. 2009, 20, 177–188. [Google Scholar] [CrossRef]
- Delgado, O.; Ruiz, J.; Pérez, M.; Romero, J.; Ballesteros, E. Effects of fish farming on seagrass (Posidonia oceanica) in a Mediterranean bay: Seagrass decline after organic loading cessation. Ocean Acta 1999, 22, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, J.M.; Romero, J. Effects of disturbances caused by coastal constructions on spatial structure, growth dynamics and photosynthesis of the seagrass Posidonia oceanica. Mar. Pollut. Bull. 2003, 46, 1523–1533. [Google Scholar] [CrossRef]
- González-Correa, J.M.; Bayle, J.T.; Sánchez-Lizaso, T.J.L.; Valle, C.; Sánchez-Jerez, P.; Ruiz, J.M. Recovery of deep Posidonia oceanica meadows degraded by trawling. J. Exp. Mar. Biol. Ecol. 2005, 320, 65–76. [Google Scholar] [CrossRef]
- Short, F.T.; Neckles, H.A. The effects of global climate change on seagrasses. Aquat. Bot. 1999, 3–4, 169–196. [Google Scholar] [CrossRef]
- Short, F.; Polidoro, B.; Livingstone, S.R.; Carpenter, K.E.; Bandeira, S.; Bujang, J.S.; Calumpong, H.P.; Carruthers, T.J.B.; Coles, R.G.; Dennison, W.C.; et al. Extinction risk assessment of the world’s seagrass species. Biol. Conserv. 2011, 144, 1961–1971. [Google Scholar] [CrossRef]
- Short, F.T.; Coles, R.G.; Pergent-Martini, C. Global seagrass distribution. In Global Seagrass Research Methods; Short, F.T., Coles, R.G., Eds.; Elsevier Science B.V: Amsterdam, The Netherlands, 2001; pp. 5–30. [Google Scholar]
- Giannoulaki, M.; Belluscio, A.; Colloca, F.; Fraschetti, S.; Scardi, M.; Smith, C.; Panayotidis, P.; Valavanis, V.; Spedicato, M.T.; Mediterranean Sensitive Habitats (MEDISEH). Final Project Report. DG MARE Specific Contract SI2, 600741, Hellenic Centre for Marine Research. 2013. 557p. Available online: http://mareaproject.net/download/71/ (accessed on 21 October 2020).
- Effrosynidis, D.; Arampatzis, A.; Sylaios, G. Seagrass detection in the Mediterranean: A supervised learning approach. Ecol. Inf. 2018, 48, 158–170. [Google Scholar] [CrossRef]
- Boudouresque, C.F.; Bernard, G.; Pergent, G.; Shili, A.; Verlaque, M. Regression of Mediterranean seagrasses caused by natural processes and anthropogenic disturbances and stress: A critical review. Bot. Mar. 2009, 52, 395–418. [Google Scholar] [CrossRef]
- Orfanidis, S.; Papathanasiou, V.; Mittas, N.; Theodosiou, T.; Ramfos, A.; Tsiolia, S.; Kosmidou, M.; Kafas, A.; Mystikou, A.; Papadimitriou, A. Further improvement, validation, and application of CymoSkew biotic index for the ecological status assessment of the Greek coastal and transitional waters. Ecol. Indic. 2020, 118, 106727. [Google Scholar] [CrossRef]
- Weatherdon, L.; Fletcher, R.; Jones, M. Manual of Marine and Coastal Datasets of Biodiversity Importance; UNEP World Conservation Monitoring Centre: Cambridge, UK, 2015. [Google Scholar]
- UNEP-WCMC; Short, F.T. Global Distribution of Seagrasses (Version 4.0). Fourth Update to the Data Layer Used in Green and Short (2003); UN Environment World Conservation Monitoring Centre: Cambridge, UK, 2018; Available online: http://data.unep-wcmc.org/datasets/7 (accessed on 15 November 2018).
- Effrosynidis, D.; Arampatzis, A.; Sylaios, G. Seagrass and hydrographic data for the Mediterranean Sea. Data Brief 2019, 25, 104286. [Google Scholar] [CrossRef]
- Ocampo-Duque, W.; Ferré-Huguet, N.; Domingo, J.L.; Schuhmacher, M. Assessing water quality in rivers with fuzzy inference systems: A case study. Environ. Int. 2006, 32, 733–742. [Google Scholar] [CrossRef]
- Kotti, I.P.; Sylaios, G.K.; Tsihrintzis, V.A. Fuzzy logic models for BOD removal prediction in free-water surface constructed wetlands. Ecol. Eng. 2013, 51, 66–74. [Google Scholar] [CrossRef]
- Telesca, L.; Belluscio, A.; Criscoli, A.; Ardizzone, G.; Apostolaki, E.T.; Fraschetti, S.; Gristina, M.; Knittweis, L.; Martin, C.S.; Pergent, G.; et al. Seagrass meadows (Posidonia oceanica) distribution and trajectories of change. Sci. Rep. 2015, 5, 12505. [Google Scholar] [CrossRef] [Green Version]
- Brix, H.; Lyngby, J. Uptake and translocation of phosphorus in eelgrass (Zostera marina). Mar. Biol. 1985, 90, 111–116. [Google Scholar] [CrossRef]
- Hemminga, M.; Duarte, C.M. Seagrass Ecology; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Duarte, C. Seagrass depth limits. Aquat. Bot. 1991, 40, 363–377. [Google Scholar] [CrossRef]
- Barbera, C.; Tuya, F.; Boyra, A.; Sanchez-Jerez, P.; Blanch, I.; Haroun, R.J. Spatial variation in the structural parameters of Cymodocea nodosa seagrass meadows in the Canary Islands: A multi-scaled approach. Bot. Mar. 2005, 48, 122–126. [Google Scholar] [CrossRef] [Green Version]
- Olesen, B.; Enríquez, S.; Duarte, C.M.; Sand-Jensen, K. Depth-acclimation of photosynthesis, morphology and demography of Posidonia oceanica and Cymodocea nodosa in the Spanish Mediterranean Sea. Mar. Ecol. Progr. Ser. 2002, 236, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Collier, C.J.; Waycott, M. Temperature extremes reduce seagrass growth and induce mortality. Mar. Poll. Bull. 2014, 83, 483–490. [Google Scholar] [CrossRef]
- Peterson, B.J.; Rose, C.D.; Rutten, L.M.; Fourqurean, J.W. Disturbance and recovery following catastrophic grazing: Studies of a successional chronosequence in a seagrass bed. Oikos 2002, 97, 361–370. [Google Scholar] [CrossRef]
- Borum, J.; Sand-Jensen, K. Is total primary productivity in shallow coastal waters stimulated by nitrogen loading? Oikos 1996, 76, 406–410. [Google Scholar] [CrossRef]
- García, R.; Holmer, M.; Duarte, C.M.; Marbà, N. Global warming enhances sulphide stress in a key seagrass species (NW Mediterranean). Glob. Chang. Biol. 2013, 19, 3629–3639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marbà, N.; Holmer, M.; Gacia, E.; Barron, C. Seagrass Beds and Coastal Biogeochemistry. In Seagrass Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R.J., Duarte, C.M., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 135–157. [Google Scholar]
- Hemminga, M.A.; Harrison, P.G.; van Lent, F. The balance of nutrient losses and gains in seagrass meadows. Mar. Ecol. Prog. Ser. 1991, 71, 85–96. [Google Scholar] [CrossRef]
Figure 1.
Fuzzy Inference System (FIS) procedures.

Figure 2.
Trapezoidal membership functions for the fuzzy sets of Bottom Chlorophyll-a imported in the FIS.
Figure 2.
Trapezoidal membership functions for the fuzzy sets of Bottom Chlorophyll-a imported in the FIS.

Figure 3.
Trapezoidal membership functions for the fuzzy sets of Surface Temperature imported in the FIS.
Figure 3.
Trapezoidal membership functions for the fuzzy sets of Surface Temperature imported in the FIS.

Figure 4.
Triangular membership functions for the fuzzy sets of the Seagrass Family, exported from the FIS.
Figure 4.
Triangular membership functions for the fuzzy sets of the Seagrass Family, exported from the FIS.

Figure 5.
Spatial distribution of Cymodoceaceae in the Mediterranean Sea (UNEP-WCMC database).

Figure 6.
Spatial distribution of Zosteraceae in the Mediterranean Sea (UNEP-WCMC database).

Figure 7.
Box-Whisker plots of annual water salinity [psu] at the locations in which seagrass families are present in the Mediterranean Sea (inner line represents sample median, box boundaries represent the distance between the first and third quartile, error bars represent the 95% CI for the median, and open circles represent the outliers).
Figure 7.
Box-Whisker plots of annual water salinity [psu] at the locations in which seagrass families are present in the Mediterranean Sea (inner line represents sample median, box boundaries represent the distance between the first and third quartile, error bars represent the 95% CI for the median, and open circles represent the outliers).

Figure 8.
Boxplots of chlorophyll-a concentration [mg/m3] at the locations in which seagrass families are present in the Mediterranean Sea (inner line represents sample median, box boundaries represent the distance between first and third quartile, error bars represent the 95% CI for the median, and open circles represent the outliers).
Figure 8.
Boxplots of chlorophyll-a concentration [mg/m3] at the locations in which seagrass families are present in the Mediterranean Sea (inner line represents sample median, box boundaries represent the distance between first and third quartile, error bars represent the 95% CI for the median, and open circles represent the outliers).

Figure 9.
Three-dimensional surface produced by the optimum FIS, showing the environmental preferences of various seagrass families. (Yellow: Posidoniaceae; Dark Blue: Zosteraceae; Green: Cymodoceaceae; Light Blue: Hydrocharitaceae).
Figure 9.
Three-dimensional surface produced by the optimum FIS, showing the environmental preferences of various seagrass families. (Yellow: Posidoniaceae; Dark Blue: Zosteraceae; Green: Cymodoceaceae; Light Blue: Hydrocharitaceae).

Figure 10.
Seagrass tolerance by family on water temperature change from initial values. Bars represent the percentage of locations/meadows that remain unchanged under different water temperature scenarios.
Figure 10.
Seagrass tolerance by family on water temperature change from initial values. Bars represent the percentage of locations/meadows that remain unchanged under different water temperature scenarios.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Seagrass family frequency in the United Nations Environment Programme—World Conservation Monitoring Centre (UNEP-WCMC) dataset (last column presents seagrasses at species level).
Table 1.
Seagrass family frequency in the United Nations Environment Programme—World Conservation Monitoring Centre (UNEP-WCMC) dataset (last column presents seagrasses at species level).
| Seagrass Gene | Locations | Percentage | Seagrass Species |
|---|---|---|---|
| Cymodoceaceae | 1337 | 75.49% | C. nodosa |
| Zosteraceae | 187 | 10.56% | Z. noltii, Z. marina |
| Posidoniaceae | 125 | 7.07% | P. oceanica |
| Hydrocharitaceae | 94 | 5.30% | H. stipulacea |
| Ruppiaceae | 28 | 1.58% | R. maritima |
Table 2.
Hydrographic and water quality parameters retrieved from CMEMS and periods these data cover. S, B represent Surface/Bottom values.
Table 2.
Hydrographic and water quality parameters retrieved from CMEMS and periods these data cover. S, B represent Surface/Bottom values.
| Parameter [Units] | CMEMS Data Product | Period |
|---|---|---|
| Significant Wave Height [m] [S] | MEDSEA_HINDCAST_WAV_006_012 | 2006–2015 |
| Water Velocity [m/s] [S, B] | MEDSEA_REANALYSIS_PHYS_006_004 | 1987–2015 |
| Water Temperature [degC] [S, B] | MEDSEA_REANALYSIS_PHYS_006_004 | 1987–2015 |
| Salinity [psu] [S, B] | MEDSEA_REANALYSIS_PHYS_006_004 | 1987–2015 |
| Chlorophyll-a [mg/m3] [S, B] | MEDSEA_REANALYSIS_BIO_006_008 | 1999–2015 |
| Nitrate [mmol/m3] [S, B] | MEDSEA_REANALYSIS_BIO_006_008 | 1999–2015 |
| Phosphate [mmol/m3] [S, B] | MEDSEA_REANALYSIS_BIO_006_008 | 1999–2015 |
| Dissolved Oxygen [mmol/m3] [S, B] | MEDSEA_REANALYSIS_BIO_006_008 | 1999–2015 |
| Net Primary Production of Biomass Rate [mol/m3/s] [S, B] | MEDSEA_REANALYSIS_BIO_006_008 | 1999–2015 |
Table 3.
Parameters for membership functions used in the fuzzy inference system. S and B represent surface and bottom values, respectively.
Table 3.
Parameters for membership functions used in the fuzzy inference system. S and B represent surface and bottom values, respectively.
| Parameter | Very Low | Low | Medium | High | Very High | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| a = b | c | d | a | b | c | d | a | b | c | d | a | b | c | d | a | b | c = d | |
| Water Depth [m] | 0 | 10 | 20 | 10 | 20 | 30 | 40 | 30 | 40 | 80 | 100 | 80 | 100 | 120 | 130 | 120 | 130 | 150 |
| Distance from Cities [km] | 0.01 | 0.26 | 0.63 | 0.62 | 0.63 | 0.87 | 1.25 | 0.87 | 1.25 | 1.50 | 1.80 | 1.50 | 1.80 | 2.10 | 2.30 | 2.10 | 2.30 | 2.70 |
| Distance from River Mouths [km] | 0.12 | 1.00 | 2.39 | 1.00 | 2.39 | 3.30 | 4.66 | 3.30 | 4.66 | 5.57 | 6.94 | 5.57 | 6.94 | 7.84 | 8.75 | 7.84 | 8.75 | 10.00 |
| Distance from Ports [km] | 0.00 | 0.12 | 0.25 | 0.12 | 0.25 | 0.37 | 0.52 | 0.37 | 0.52 | 0.62 | 0.75 | 0.62 | 0.75 | 0.87 | 1.00 | 0.87 | 1.00 | 1.20 |
| Distance from Coast [km] | 0.00 | 0.13 | 0.34 | 0.13 | 0.34 | 0.48 | 0.68 | 0.48 | 0.68 | 0.82 | 1.00 | 0.82 | 1.00 | 1.17 | 1.31 | 1.17 | 1.31 | 1.50 |
| Temperature [S] [degC] | 15.0 | 15.5 | 16.5 | 15.5 | 16.5 | 17.5 | 18.0 | 17.5 | 18.0 | 19.0 | 19.5 | 19.0 | 19.5 | 20.5 | 21.0 | 20.5 | 21.0 | 23.0 |
| Temperature [B] [degC] | 11.0 | 12.0 | 13.0 | 12.0 | 13.0 | 14.0 | 15.0 | 14.0 | 15.0 | 17.0 | 18.0 | 17.0 | 18.0 | 20.0 | 21.0 | 20.0 | 21.0 | 22.0 |
| Salinity [B] [psu] | 36.0 | 36.3 | 36.7 | 36.3 | 36.7 | 37.0 | 37.4 | 37.0 | 37.4 | 38.1 | 38.5 | 38.1 | 38.5 | 39.2 | 39.6 | 39.2 | 39.6 | 40.0 |
| Nitrates [S] [mmol/m3] | 0.0 | 1.5 | 3.5 | 1.5 | 3.5 | 5.0 | 7.0 | 5.0 | 7.0 | 9.0 | 11.0 | 9.0 | 11.0 | 13.0 | 15.0 | 13.0 | 15.0 | 16.0 |
| Nitrates [B] [mmol/m3] | 0.0 | 1.4 | 3.4 | 1.4 | 3.4 | 4.8 | 6.9 | 4.8 | 6.9 | 8.2 | 10.3 | 8.2 | 10.3 | 11.7 | 13.1 | 11.7 | 13.1 | 15.0 |
| Phosphates [B] [mmol/m3] | 0.0 | 0.04 | 0.08 | 0.04 | 0.08 | 0.11 | 0.15 | 0.11 | 0.15 | 0.19 | 0.23 | 0.19 | 0.23 | 0.26 | 0.30 | 0.26 | 0.30 | 0.38 |
| DO [S] [mmol/m3] | 145 | 156 | 167 | 156 | 167 | 178 | 189 | 178 | 189 | 200 | 211 | 200 | 211 | 222 | 233 | 222 | 233 | 255 |
| Chl-a [B] [mg/m3] | 0.00 | 0.06 | 0.12 | 0.06 | 0.12 | 0.18 | 0.24 | 0.18 | 0.24 | 0.30 | 0.36 | 0.30 | 0.36 | 0.42 | 0.48 | 0.42 | 0.48 | 0.60 |
Table 4.
Indicative fuzzy rules in initial FIS test, where VL represents locations with values belonging to the Very Low fuzzy set, L in the Low, M in the Medium, and H in the High fuzzy set.
Table 4.
Indicative fuzzy rules in initial FIS test, where VL represents locations with values belonging to the Very Low fuzzy set, L in the Low, M in the Medium, and H in the High fuzzy set.
| No | Distance from Cities | Phosphates [S] | Salinity [B] | Chl-a [B] | Seagrass Family |
|---|---|---|---|---|---|
| 1 | M | VL | M | L | Zosteraceae |
| 2 | L | VL | M | M | Zosteraceae |
| 3 | VL | H | M | VL | Zosteraceae |
| 4 | VL | VL | M | H | Zosteraceae |
| 5 | VL | VL | M | M | Zosteraceae |
| 6 | L | L | H | VL | Hydrocharitaceae |
| 7 | M | L | H | VL | Hydrocharitaceae |
| 8 | VL | L | H | VL | Hydrocharitaceae |
| 9 | L | VL | M | L | Cymodoceaceae |
| 10 | VL | VL | M | L | Cymodoceaceae |
| 11 | L | L | M | L | Cymodoceaceae |
| 12 | L | VL | H | L | Posidoniaceae |
| 13 | M | M | H | VL | Posidoniaceae |
Table 5.
Indicative fuzzy rules in the optimum FIS, where VL represents locations with values belonging to the Very Low fuzzy set, L in the Low, M in the Medium, H in the High, and VH in the Very High fuzzy set.
Table 5.
Indicative fuzzy rules in the optimum FIS, where VL represents locations with values belonging to the Very Low fuzzy set, L in the Low, M in the Medium, H in the High, and VH in the Very High fuzzy set.
| No | Water Depth | Chl-a [B] | Temperature [S] | Nitrates [S] | Seagrass Family |
|---|---|---|---|---|---|
| 1 | VL | M | L | VL | Zosteraceae |
| 2 | VL | H | L | L | Zosteraceae |
| 3 | VL | L | M | VL | Zosteraceae |
| 4 | VL | M | L | L | Zosteraceae |
| 5 | H | VL | M | VL | Hydrocharitaceae |
| 6 | H | VL | VH | VL | Hydrocharitaceae |
| 7 | L | VL | VH | VL | Hydrocharitaceae |
| 8 | M | VL | H | VL | Hydrocharitaceae |
| 9 | L | L | H | VL | Cymodoceaceae |
| 10 | VL | L | H | VL | Cymodoceaceae |
| 11 | L | M | H | VL | Cymodoceaceae |
| 12 | L | VL | H | VL | Cymodoceaceae |
| 13 | L | L | L | VL | Posidoniaceae |
| 14 | VL | L | L | VL | Posidoniaceae |
| 15 | M | M | M | VL | Posidoniaceae |
| 16 | M | H | H | VL | Posidoniaceae |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Papaki, D.; Kokkos, N.; Sylaios, G. A Fuzzy Inference System for Seagrass Distribution Modeling in the Mediterranean Sea: A First Approach. Water 2020, 12, 2949. https://doi.org/10.3390/w12102949
AMA Style
Papaki D, Kokkos N, Sylaios G. A Fuzzy Inference System for Seagrass Distribution Modeling in the Mediterranean Sea: A First Approach. Water. 2020; 12(10):2949. https://doi.org/10.3390/w12102949
Chicago/Turabian StylePapaki, Dimitra, Nikolaos Kokkos, and Georgios Sylaios. 2020. "A Fuzzy Inference System for Seagrass Distribution Modeling in the Mediterranean Sea: A First Approach" Water 12, no. 10: 2949. https://doi.org/10.3390/w12102949
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.